This is the old United Nations University website. Visit the new site at http://unu.edu
The floristic composition of the caatinga in Campina Grande is shown in table 12.1. Mimosa tenuiflora and Caesalpinia pyramidalis predominate, with Pithecellobium foliorosum and Aspidosperma pyrifolium also present. In the shrub layer, Croton sonderiana, Sida cordifolia, and Croton campestris were the dominant species. Observation indicated that the bare sites produced by disturbance were covered first by Sida cordifolia and Croton campestris. These were then replaced by Mimosa tenuiflora and Caesalpinia pyramidalis. The final stage of succession of this area appears to be Aspidosperma pyrifolium, be cause the trees are of big stature, with erect stems, emerging above the other tree species.
Table 12.1 Floristic composition of the caatinga at the vicinity of Campina Grande (quadrat area 10 m x 10 m; 1st and 2nd layers above and less than 1 m in height)
| Species | 1st layer | 2nd layer | |
| Mean DBH (cm) | No. of trunks | No. of individuals | |
| Caesalpinia pyramidalis | 4.4 | 12 | 7 |
| Pithecellobium foliorosum | 4.2 | 4 | 0 |
| Mimosa hostilis | 4.0 | 28 | 3 |
| Croton sonderiana | 3.3 | 4 | 13 |
| Aspidosperma pyrifolium | 2.7 | 4 | 3 |
| Encholrium spectabile | - | - | 8 |
| Spondias tuberosum | - | - | 1 |
| Croton species | - | - | 1 |
| Pilosocereus peutedrophorus | - | - | 1 |
| Unidentified species 1 | - | - | 2 |
| Unidentified species 2 | - | - | 2 |
Table 12.2 Quandiative caracteristics of caatinga stand at the experimental sta tion,
| Campina Grade | |
| Number of species per 100 m² | |
| Tree layer | 7 |
| Herb layer | 13 |
| Number of trees per 100 m² | 39 |
| Mean DBH (cm) | 4 |
| Above-ground biomass (kg per 100 m²) | |
| Trunks and branches | 246 |
| Leaves | 29 |
| Total | 275 |
| Herbs | 1 9 |
| Gross total | 294 |
The Caesalpinia pyramidalis stand included 11 species and 52 individuals per 100 m². The height of the tree layer was about 5 m. The density and mean DBH of the trees were 39-52 individuals per 100 m² and 3.7-4 cm. The aboveground phytomass of the stand was 294 kg per 100 m², including 246 kg of trunks and branches, 29 kg of leaves and 19 kg of herbs and grasses (table 12.2).
The relationship between mean DBH and number of trees per 100 m² (tree density) is shown in figure 12.2, which was obtained by the survey of 9 stands of caatinga. The relationship shown in this figure is described as follows:
N= 217 exp ( - 0.42 D) (1)
where N and D are tree density and mean DBH. This equation suggests that the number of trees per unit area decreased exponentially with increment of mean DBH.
The relationship between DBH (cm) and weight mass of trees (W:kg), trunks and branches (Wt:kg) and leaves (Wl:kg) is shown in figure 12.3. These relationships are as follows:
W = 0.226 D2.274 (2)
Wt = 0.206D2.273 (3)
Wl = 0 .018D2.369 (4)
After measuring the diameter of stem, we are able to estimate the tree weight from equation (2). The tree weights of Caesalpinia pyramidalis and Aspidosperma pyrifolium were 13.3 kg and 22.8 kg in estimated, and 14.5 kg and 23.0 kg in measured values, respectively (Hayashi, 1986).
The area of the annual ring formed in the stem of Pithecellobium foliorosum was computed for each year from 1975 to 1985. Based on the area of annual ring (s:cm²), I obtained the stem diameter (d:cm) using the following equation:
![]() (5)
(5)
The DBH was obtained by adding bark thickness to the stem diameter (d). The growth of DBH obtained above is shown in figure 12.4. for Pithecellobium foliorosum.
The growth curve of DBH of P. foliorosum was described as
D (t) = 9/{1 + 1.04 exp [ - 0.14 t]} (6)
where D (t) is DBH at year t, t is year from 1976 (1976 is 0 year). As shown by this figure, the logistic curve obtained by the least-squares method describes the growth of this tree. According to the equation, the maximum DBH and relative growth rate are 9 cm and 0.14 per year.
In a previous paper (Hayashi, 1986), I reported the same relationship for Caesalpinia pyramidalis and Aspidosperma pyrifolium, which are the dominant species of the caatinga. These equations were as follows:
D (t) = 7.7/{1 + 1.16 exp ( - 0.098 t)} (7)
(Caesalpinia pyramidalis: from 1963 to 1984)
D (t) = 12.3/{1 + 2.33 exp ( - 0.067 t)} (8)
(Aspidosperma pyrifolium: from 1963 to 1984)
Assuming that the trees in the caatinga grow in accordance with equations (6), (7), and (8), based on a computer simulation, I predicted a change of DBH histogram of a caatinga stand from 1984 to 1989. The DBHs of trees measured in 1984 in Campina Grande were used in the simulation. The changes in the DBH histogram produced by computer simulation are given in figure 12.5. According to these results, the mean DBH changed from 4.1 cm in 1984 (measured value) to 6.4 cm in 1989.
The number of trees per unit area (predicted value) is expected to decrease according to equation (1) from 39 individuals per 100m² in 1984 to 16 individuals in 1989. During this period, the tree mortality in the stand from self-thinning is expected to be 3 from 1984 to 1985, 6 from 1985 to 1986, 7 from 1986 to 1987, 4 from 1987 to 1988 and 3 from 1988 to 1989. A total of 23 trees from the stand are expected to die during the five years.
Table 12.3 Carbon and nitrogen of the plants of the doniinant species in caatinga
| Species | Carbon (%) | Nitrogen (%) | C:N |
| Mimosa hostilis | |||
| Wood | 46.0 | 1.0 | 49.7 |
| Bark | 52.1 | 1.7 | 31.2 |
| Leaf | 43.1 | 2.4 | 18.0 |
| Caesalpinia pyramidalis | |||
| Wood | 47.1 | 1.0 | 49.5 |
| Bark | 45.5 | 1.2 | 37.6 |
| Leaf | 46.1 | 2.5 | 19.0 |
| Aspidosperma pyrifolium | |||
| Wood | 47.4 | 1.1 | 43.1 |
| Leaf | 42.2 | 2.7 | 15.7 |
| Panicum trichoides | |||
| Whole plant | 44.2 | 2.8 | 15.7 |
| Encholrium spectabile | |||
| Whole plant | 46.1 | 1.7 | 26.5 |
| Herhaceons. plants | 41.5 | 2.5 | 16.6 |
The mass of leaves, trunks, and branches from dead trees was estimated, using equations (2), (3), and (4). Based on the DBH shown in figure 12.5 and using the above equation, it is possible to estimate the total mass of litter for each year. Substituting the DBH of dead trees (which are assumed to be the smaller trees within the stand), with equation (3), the mass of trunks and branches from the dead trees were estimated for each year. The mass of fallen leaves at the end of rainy season was also estimated using equation (4). Thus, the total mass of litter supplied to the stand is expected to be 280 kg (164 kg of leaves and 116 kg of trunks and branches) per 100m² during the five years from 1985 to 1989 (Hayashi, 1986, 1988).
The carbon and nitrogen content of the plants are given in table 12.3. Nitrogen content was 2.4-2.8 per cent in the leaves, 1.0-1.1 per cent in the wood and 1.2-1.7 per cent in the bark. The C:N ratios were 43.1-49.7 for the wood and 15.7-19.0 for the leaves, respectively. Uhl et al. (1982) reported that the leaves of tree shoots from the Amazonian caatinga contained 0.78 per cent nitrogen, which is one third of the nitrogen content in the leaves of caatinga trees in North-East Brazil. The plant carbon contents were: wood 46-47 per cent, leaves 42-46 per cent and bark 45-52 per cent. Multiplying 2.4 per cent and 1.05 per cent by total mass of leaf and trunk litter, 5.26 kg/100m² of nitrogen is expected to be supplied to the stand in the form of plant litter during a five-year period. The carbon supplied to ground surface by the litter is expected to be 126.8 kg during the same period, which is equivalent to 466 kg of carbon dioxide (table 12.4). This suggests that the vegetation absorbs the 466 kg per 100m² of carbon dioxide from the atmosphere during the fiveyear period.
Table 12.4 Amount of plant litters and of carbon and nitrogen in the litters produced by the caatinga stand for 5 years (kg/100m²)
| Litter | Carbon | Nitrogen | Carbon dioxide | |
| Leaf | 164 | 71.8 | 4.10 | 264 |
| Trunk and branch | 116 | 55.0 | 1.16 | 202 |
| Total | 280 | 126.8 | 5.26 | 466 |
Table 12.5 Soil texture of the caatinga stands under different human impacts in the vicinity of Campina Grande
| Soil depth (cm) | Content in percentage | ||||
| Clay | Silt | Fine sand | Coarse sand | ||
| Site 1 | |||||
| 0-5 | 14.0 | 10.2 | 71.3 | 4.5 | SL |
| 6-10 | 19.4 | 11.4 | 62.0 | 6.9 | SCL |
| 11 -15 | 23.7 | 12.8 | 57.3 | 6.2 | SCL |
| 16-20 | 25.1 | 11.0 | 58.5 | 4.9 | SC |
| 21-25 | 23.5 | 12.1 | 58.0 | 6.4 | SCL |
| Site 2 | |||||
| 0-5 | 19.4 | 9.1 | 54.1 | 17.4 | SCL |
| Site 3 | |||||
| 0-5 | 5.0 | 9.3 | 71.0 | 14.7 | SL |
Site 1: developed stand of caatinga;
site 2: disturbed stand of caatinga; 11 km south from C. Grander
site 3: heavily disturbed stand in the vicinity of C. Grande.
The texture of caatinga soils is given in table 12.5. The fine sand decreases with depth, although the coarse sand content remains constant. By contrast, the clay content increased with soil depth. Surface soils of disturbed and severely disturbed sites contained a larger proportion of coarse sand than that of developed stands.
Soil nitrogen and carbon concentrations are given in table 12.6. The top 20 cm of caatinga soil contained 0.01-0.14 per cent of nitrogen and 0.39-1.65 per cent of carbon. The soil nitrogen is very low comparable to B horizon for soils of well-developed temperate forest. Grove et al. (1986) reported that the surface soil of a Eucalyptus marginata forest in south-western Australia, which is similar to caatinga in physiognomy, contained 0.14 per cent of nitrogen. After burning, the soil nitrogen increased to 0.17 per cent. The nitrogen content before the fire was similar to the caatinga soil.
Table 12.6 Carbon (C) and nitrogen (N) content (%) of the soils of caatingas under different human impacts at the vicinity of Campina Grande
| Soil depth (cm) | Site 1 | Site 2 | Site 3 | |||
| C | N | C | N | C | N | |
| 0-5 | 1.14 | 0.02 | 1.65 | 0.14 | 0.67 | 0.05 |
| 6-10 | 0.56 | 0.02 | 0.75 | 0.07 | - | - |
| 11-15 | 0.43 | 0.01 | 0.46 | 0.05 | - | - |
| 16-20 | 0.39 | 0.01 | 0.43 | 0.04 | - | - |
Site 1: developed stand of caatinga;
Site 2: disturbed stand of caatinga 11 km south from C. Grander
Site 3: heavily disturbed stand of caatinga at the vicinity of C.
Grande.
The flora of the semi-arid region of North-East Brazil was described by Luetzelburg (1922-23). According to him, the dominant species in the caatinga were Caesalpinia ferrea, Caesalpinia pyramidalis, Caesalpinia echinata, Aspidosperma pyrifolium, Spondias tuberosa, Magonia pubescens, Hymenaea martiana, Mimosa verrucosa, Mimosa tenuiflora, Cereus squamosus, Cereus jamacaru, and Pilocereus gounellei. Recently, Gomez (1981) studied the vegetation of Cariris Velhos, Paraíba state, and reported that the dominant species in that region were Caesalpinia pyramidalis, Mimosa tenuiflora, Croton sonderianus, Combretum leprosum, Aspidosperma pyrifolium, Jatropha pholiana, Spondias tuberosa, Pithecellobium foliorosum, and Pilocereus gounellei. These floristic compositions are similar to my results in Campina Grande and Patos.
According to Whittaker (1970), the average biomass of shrubland formation is in the range of 2 to 20 kg per m², which includes the value of 2.94 kg m², thus bracketing our value of 2.94 kg m² obtained from stands used for firewood and ranching. The phytomass of the stands varies from site to site according to the intensity of human activities.
The area has been degraded by varying human impacts such as insufficient fallowing period within the shifting cultivation; charcoal production; felling wood for fuel used in brick production; demand for fuel wood by the bakeries in (Campina Grande and Patos; and fence construction to enclose goats (fig. 12.6).
According to Saito et al. (1988), productivity of cultivated crops for 1986 in the município of Patos was 1,050 kg per 9,683 ha, for perennial and annual cotton; 44 kg per 2,299 ha, for maccather bean; 600 kg per 1,962 ha, for maize; and 1,200 kg per 60 ha, for rice. Livestock figures in Patos in 1980 were 14,274 head of cattle, 1,762 head of sheep, and 293 head of horses for the total area of 59,499 ha. Nishizawa and Pinto (1988) reported that during 1981, 16,253 tons of charcoal and 1,569,612m³ of firewood were consumed in the region, including Seridó Paraibano, Piemonte da Borborema, Depressão do Alto Piranhas, Cariris Velhos, and Agreste da Borborema. In 1981, 1,890m³ of timber was produced for house and fence construction. Saito et al. (1988) noted that a single family consumes 100 kg of firewood in a week (five tons per year). Increasing demand for firewood by the bakeries in Patos was also reported by Nishizawa and Pinto (1988).
These human activities have resulted in a lot of bare ground within NorthEast Brazil, thus promoting land degradation. Once the vege tation has been removed, tropical rains create a sandy surface soil and vegetation recovery, including seed germination and seedling establishment, is limited by the severe condition of the soil surface (Hayashi 1981, 1988). We should, with proper use of the land, conserve plant cover to protect the soil against erosion. It is necessary, therefore, that the production of livestock, crops, and fuels be managed within the capabilities of the natural biological potential for the area.
Gomez, M.A.F. 1981. Padrães de caatinga nos Cariris Velhos, Paraíba. Ceará, Fortaleza.
Grove, T.S., A.M. O'Connell, and G.M. Dimmonk. 1986. "Nutrient changes in surface soil after an intense fire in jarrah (Eucalyptus marginata Donn ex Sm.) forest." Australian Journal of Ecology 11: 303-17.
Hayashi, I. 1981. "Plant communities and their environments in the caatinga of Northeast Brazil." Latin American Studies 2: 66-79.
. 1986. "Vegetation and soils of humid and semi-arid regions in Northeast Brazil." Latin American Studies 8: 49-62.
. 1988. "Changing aspect of caatinga vegetation in semi-arid region, Northeast Brazil." Latin American Studies 10: 61-76.
Luetzelburg, P. von. 1922-23. Estudo botônico do Nordeste, vols. 2, 3. Rio de Janeiro.
Ministério da Agricultura e Ministério do Interior. 1971. Mapa exploratorio-reconhe-cimento de solos: Estado da Paraíba (1:500,000). FIBGE, Rio de Janeiro
Nishizawa, T. 1976. "Some characteristics of rainfall in the Northeast of Brazil." Tokyo Geography Papers XX: 53-61.
Nishizawa, T., and M.M.V. Pinto. 1988. "Recent changes in firewood and charcoal production and their affects on deforestation." Latin American Studies 10: 122-31.
Rizzini, C.T., and M.M. Pinto. 1964. "Areas climático-vegetacionais do Brasil, segundo os métodos de Thornthwaite e Mather." Revista Brasileira de Geografia 26 (4): 523-47.
Saito, I., H. Maruyama, and K.D. Muller. 1988. "A comparative study of land use between the Campo Alegre in the sertão and Sitio Açude de Pedra in the agreste, Paraíba, Northeast Brazil." Latin American Studies 10: 78-99.
Uhl, C., C. Jordan, K. Clark, H. Clark, and R. Herrera. 1982. "Ecosystem recovery in Amazon caatinga forest after cutting, cutting and burning, and bulldozer clearing treatments." OIKOS 38: 313-20.
Whittaker, R.H. 1970. Communities and ecosystems. Collier-Macmillan, London.
Characteristics and utilization of tree species in the semi-arid woodland of north-east Brazil
1
Introduction
2 Bioclimatic divisions of north-east Brazil and
the floristic composition of the caatinga stand
3 Response and stress tolerance of caatinga
trees to various water conditions
4 Utilization of the caatinga trees
5 Deforestation associated with increased
firewood consumption and charcoal production
6
Conclusions and a proposal
Acknowledgements
Appendix
References
Toshie Nishizawa, Akio Tsuchiya, and Maria Magdalena Vieira Pinto
A large expanse of deciduous thorny woodland, caatinga covers the semiarid interior of North-East Brazil. The annual rainfall, which occurs mostly between December and April, varies from 400 to 800 mm. Owing to the dryness of the climate, aridisols, alfisols, and entisols are dominant. The capacity of these soils to hold water is limited by their shallowness and low organic matter content.
The Tupi word caatinga, literally "white forest," is descriptive of the open vegetation, the bleached appearance of the light-coloured trees and soils found during the dry season. With the arrival of the rainy seasons the caatinga changes completely: a paradise of lush vegetation bursts forth, birds sing, and butterflies flutter around.
Subsistence agriculture and open-range cattle grazing were introduced to the semi-arid area in the seventeenth century. Although crop farming provides greater economic benefits, it is limited mostly to the floodplains of intermittent rivers, and the moist bottoms of reservoirs (açudes), uncovered as the level of impounded water drops.
Agriculture gradually changed during the past two hundred years with the introduction of cash crops such as cotton and agave. However, subsistence farming of the moist areas still continues, especially the practice of interplanting crops such as maize, two or three kinds of beans, and squash. Normally, the following sequence occurs annually in the semi-arid interior, the sertão. During the dry season livestock graze in the fields. At the end of the rainless season fields are prepared for planting, and if the rains arrive on time and in adequate amounts the crops grow sufficiently to produce a harvest. This pattern of agricultural land use is repeated annually. In the past, after several years the cultivated fields were left to fallow and a second growth, or capoeira. However, recent population increases and economic pressures have modified this pattern of land use. Fallow periods are shortened or eliminated. The result of this overuse is accelerated erosion and depletion of soils, as well as the invasion of less desirable plant species and a general degradation of the ecosystem.
Livestock consists of cattle, sheep, and goats. Which of these animals are chosen is determined, not only by the variable forage production due to the interannual fluctuation of rainfall, but also by the mix of the preferred forage species. Moreover, the choice of livestock types is influenced strongly by the investment which individual farmers are willing - or financially able - to risk: cattle and sheep require greater capital than goats.
In addition to forage, the caatinga provides farmers with construction materials, fuelwood, charcoal, honey, fruits, fibres, and medicinal plants. Furthermore, trees are removed from the caatinga for commercial fuel in bakeries and in both nearby and distant brick and tile factories. Thus the caatinga provides an environment for multiple land use under traditional forest management.
This paper will discuss: ¹ the strategies for adaptation of caatinga trees to the severe natural conditions of the semi-arid interior; and ² possible improved utilisation of the caatinga stand.
2 Bioclimatic divisions of north-east Brazil and the floristic composition of the caatinga stand
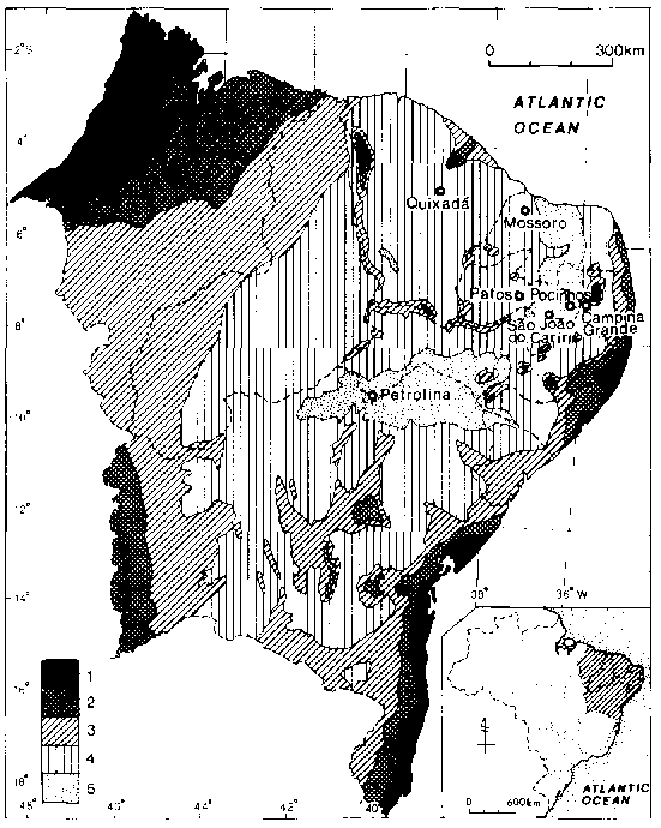
Two major subdivisions are commonly recognized in the Brazilian NorthEast, or Nordeste. There is the western Nordeste (roughly the states of Maranhão and Piaui), often called the Meio Norte ("Half North") because its climate and vegetation are transitional in respect to those of Amazonia (the Norte). Then there is the eastern Nordeste, which will be the focus of this paper. This region is, in turn, divided longitudinally, starting from the humid coastal strip (zona da mata), through a transitional belt (agreste), to the semi-arid backlands(sertão). Golfairi and Caser (1977) divide the Nordeste into five bioclimatic regions, (table 13.1 and fig. 13.1. The pocket-shaped area which includes Region 4 and Region 5, except for small but ecologically and culturally significant enclaves, was originally covered by caatinga vegetation. These regions lie in areas of less than 1,000 mm of annual precipitation and annual water deficits greater than 500 mm. However, the amount of 300 mm/m as a soil's water-holding capacity, used by Golfairi and Caser for the calculation of water deficit, is much too great for the shallow soils in the caatinga.
Table 13.1 Climatic indices for biorlimatic division (after Golfairi and Caser, 1977)
| Region | Mean
annual air temp. (°C) |
Annual
rain- fall (mm) |
Annual
water deficit (mm) |
Humidity index |
Relative
humidity (%) |
| 1 | 20 ~ 27 | 1,500 ~ 1,350 | 0 ~ 100 | + 20 ~ + 100 | 78 ~ 90 |
| 2 | 20 ~ 27 | 1,000 ~ 1,700 | 50 ~ 300 | 0 ~ + 20 | 70 ~ 80 |
| 3 | 20~27 | 700~1,300 | 200~ 600 | 0~ -33 | 65~76 |
| 4 | 21~28 | 500~1,000 | 500~1,000 | -33~ -66 | 60~70 |
| 5 | 21~28 | 250~ 550 | 800~1,300 | -66~ -100 | 45~65 |
From the correlation between seasonal discharge and water surplus in three drainage basins of the semi-arid backland, Tsuchiya (1990) calculated the value of 50 mm/m as the soil water-holding capacity of the caatinga region from 1968 to 1987. Table 13.2 shows annual water storage and deficit, using a water-holding capacity of 50 mm/m at three stations.
Although Campina Grande and Patos, both in Paraíba State, are located in the same Region 4 defined by Golfairi and Caser, annual water deficits differ by about 600 mm. While Patos and Petrolina, Pernambuco State, are located in different regions, their annual water deficits are almost the same, as are their figures for the seasonal marches of the water balance.
The dominant species and the floristic composition of the caatinga are affected initially by natural conditions such as water balance and soil properties, but also by human activity. The magnitude of human impact on the caatinga has increased rapidly in recent years, owing to population and economic pressure, which results in greater scarcity of mature stands.
Trees of the caatinga stand with heights greater than 1 m are described by their floristic composition for five sites, but trees with heights smaller than 1 m are included at Campo Alegre I (table13.3). Campo Alegre, located 15 km north of Patos, is in Region 4; Pocinhos is in Region 5, as are Nogueira and Varginha, both 40 km east of Petrolina.
Figure 13.1 Bioclimatic regions of North-East Brazil. (After Golfairi and Caser, 1977.)
The floristic composition of Campo Alegre I is characterized by a twoyear regrowth of caatinga. At Campo Alegre II, cattle grazing occurs with selective cutting of trees for charcoal production. Nogueira has low-density cattle grazing, whereas in Varginha only goats and sheep browse. Malva is by far the dominant species in Campo Alegre I, while marmeleiro and jurema-preta prevail in Campo Ale are II. For Pocinhos, the jurema-preta species dominates, followed by catingueira and marmeleiro. Generally, jurema-preta is one of the dominant species at disturbed sites (Hardesty et al., 1988). Catingueira is the dominant species in both Nogueira and Varginha. Three sites have five species over one metre high, with jurema-preta and/or catingueira dominant. However, young jurema-preta never appear with mature catingueira, while young catingueira trees occur with mature jurema-preta and with other trees. Thus, it can be concluded that, following clear-cutting species such as malva and/or marmeleiro, jurema-preta, and catingueira dominate at different phases of caatinga regrowth. (For local and scientific names of caatinga trees, see Appendix, p. 298.)
Table 13.2 Means of annual water storage and water deficit for twenty years from 1968 to 1987
| Station | Water storage (mm) | Water deficit (mm) |
| Campina Grande | 181 | 443 |
| Patos | 118 | 1,093 |
| Petrolina | 62 | 1,047 |
Table 13.3 Flonstic composition of the caatinga stand at five sites
| Sites | No. of stems | (%) |
| (1) Campo Alegre I (10m x 10m) | ||
| Malva | 506 | 92 |
| Pinhão | 25 | 5 |
| Marmeleiro | 10 | 2 |
| Jurema-preta | 3 | 1 |
| (2) Campo Alegre II (l0m x 10m) | ||
| Marmeleiro | 79 | 72 |
| Jurema-preta | 23 | 21 |
| Catingueira | 6 | 6 |
| Mofumbo | 1 | 1 |
| (3) Pocinhos (10m x 10m) | ||
| Jurema-preta | 23 | 52 |
| Catingueira | 10 | 23 |
| Marrneleiro | 7 | 16 |
| Pinhão | 2 | 5 |
| Cipo | 1 | 2 |
| Mestracio | 1 | 2 |
| (4) Varginha (20m x 20m) | ||
| Catingueira | 34 | 57 |
| Jurema-preta | 20 | 33 |
| Faveleira | 3 | 5 |
| Pinhão | 2 | 3 |
| Umbuzeiro | 1 | 2 |
| (5) Nogueira (10m x 10m) | ||
| Catingueira | 18 | 58 |
| Pinhão | 7 | 23 |
| Jurema-preta | 3 | 10 |
| Burra-leiteira | 2 | 6 |
| Faveleira | 1 | 3 |
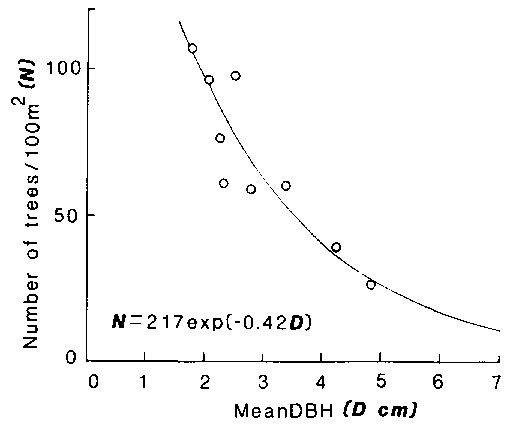{kind=link}
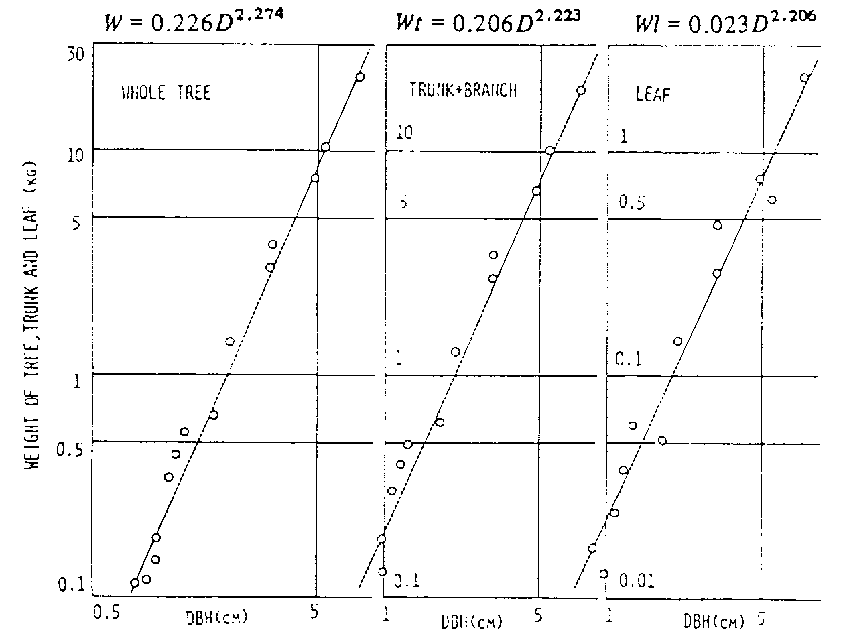{kind=link}
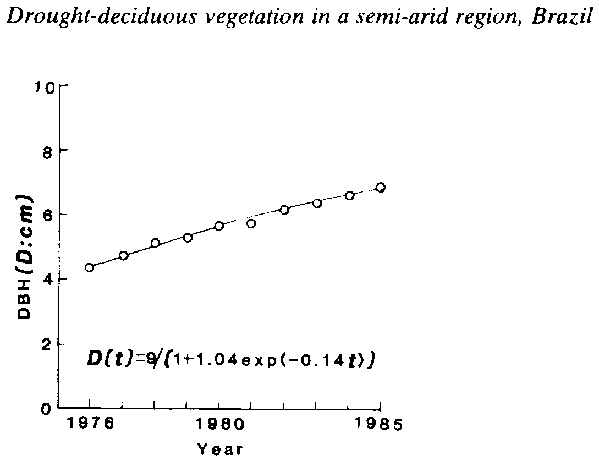{kind=link}
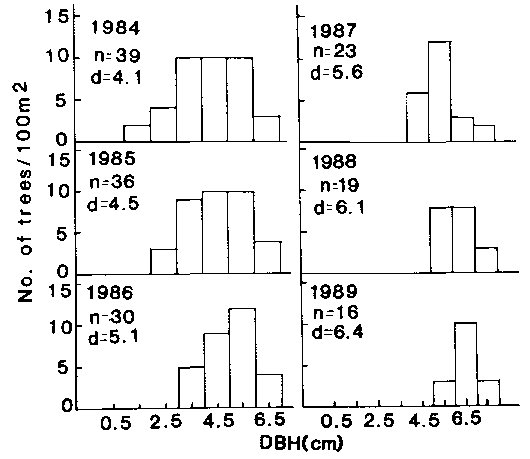{kind=link}
{kind=link}