This is the old United Nations University website. Visit the new site at http://unu.edu
Food losses through energy transfer from cereal grains to stored-product insects
R.N. Sinha
Agriculture Canada, Research Station, Winnipeg,
Manitoba, Canada
A sound strategy for the reduction of food losses during post-harvest storage requires the development of standardized quantitative criteria to measure losses that are characteristic of geographical location, climate, and food type. With advancement in food-storage research and relatively predictable food crop production, it is reasonable to expect that production estimates based primarily on the genetic potential of crops may soon be matched with almost equally reliable post-harvest food loss estimates. To determine and predict food losses accurately, in standardized units of measurement, basic research data must be known on the potential of each pest species to inflict loss on each type of food crop stored under optimal and suboptimal environmental conditions. The extent of damage should be expressed in terms of one or more acceptable scientific criteria. Various criteria have been used to measure food loss caused by the presence or action of insects, mites, moulds, rodents, and birds. Some of these criteria are: weight loss, quality loss as expressed by degradation of the nutritional and aesthetic value of a food contamination by animal excrete or mycotoxins, monetary loss (e.g., insurance), loss of goodwill of importers, and seed germination loss (1, 2). Formulation of a loss measurement criterion that will be scientifically sound and universally acceptable is necessary for the budgeting of food resources and developing rational storage and distribution systems for food grain.
Energy in the system
By following a fundamental principle of thermodynamics -i.e., energy is neither created nor destroyed, but can be transformed -energy flow from food grain to pest organisms, such as insects, can be accurately measured and used as a scientifically defensible criterion for assessing food loss. Considering that plant seeds have genetically determined limited caloric energy, the actual and potential caloric energy consumed by individuals of an insect species can be determined by measuring the exact amount of energy removed from individual seeds. Food energy flows from crop plant to crop seed, to dormant stored seed, and then to the body tissues of the insects feeding on stored grain. Interference of this flow can reduce food loss.
If we wish to interfere with the process of spoilage of stored food through energy transfer - i.e., from cereal plant seed in a warehouse to mould, insect, or rodent body tissue and their metabolic by-products-we may do it on a longer term basis if we consider a grain mass as an ecosystem in the same way an ecologist regards a pond as an ecosystem. Odum (3) defines an ecosystem as any area of nature that includes living organisms and non-living materials interacting to produce an exchange of materials between the living and non-living parts of the system. An ecosystem, which is powered either directly or indirectly by solar energy, is an arbitrarily defined unit in which there is a distinct pattern of energy flow and chemical cycling (4).
Solar energy reacts with chemicals from the environment and forms the plant tissues through the photosynthetic process. Green plants are called "producers" because they produce organic biomass using the energy of the sun to transform assimilated inorganic substances into organic material; some of the latter is used for metabolism and maintenance of the plant. A portion of the energy released as heat and chemical constituents is returned to the environment to re-enter the nutrient cycle. The material left after metabolism and assimilation is called biomass or actual growth. The biomass energy disintegrates through death or is transformed into the biomass (growth) or metabolic activity of another living organism, the consumer that eats the plant. The assemblage of plant and animal forms is an ecosystem containing a producer, several levels of consumers, and decomposers that include microflora that are involved in the disintegration of dead plant and animal bodies (5).
Granaries as ecosystems
An ecosystem is composed of species, populations, and communities of plants and animals. A population is a group of organisms of the same species occupying a particular space at a particular time. A community is composed of groups of coexisting, interdependent populations of plants and animals in a given area. The groups may be of different sizes and degrees of integration, e.g., the insect community in a silo filled with corn. In an ecosystem, communities of micro-flora, mites, insects, birds, and rodents may operate together and respond to the changing abiotic environment dominated by the interplay of temperature and the relative humidity of the air (5). Even if only together for a short time, the organisms achieve some sort of functional stability. An ecosystem is also regarded as an assembly of interlocking life systems.
A granary containing a grain crop such as wheat, barley, or corn is a short-lived, man-made ecosystem with a nonregenerating supply of energy. The solar energy trapped as carbohydrates in chlorophyll-bearing cereal plants that are a part of the pre-harvest agro-ecosystem is accumulated in seeds along with mineral nutrients captured by roots from the soil, and together they are used to maintain the post-harvest stored-grain ecosystem (fig. 1).
Despite its temporary functional stability, a stored-grain ecosystem is inherently unstable because of disturbances frequently caused by climate and man. Characteristically, it contains: a handful of species with minute to small individuals with morphologically simple, short life spans and facultative interrelations, but with high growth and reproductive rates; short food chains; and a relatively high concentration of energy per unit of biomass. Unlike mature and stable ecosystems, such as a tropical forest, the interrelations among various biotic and abiotic variables in this type of immature ecosystem are often vague and too transitory to quantify and define (6). Nevertheless, the bulk-grain ecosystem represents valuable food resources, and despite voracious insects and dangerous decay-causing and toxin-producing micro-organisms that inhabit such an ecosystem, its proper management is crucial to the survival of the human race.
Only a few attempts have been made to learn the operative mechanisms of both the plant and animal communities within these ecosystems (7,8,9). To my knowledge, no total energy budget study (including most measurable biotic and abiotic components) of any stored-grain ecosystem has ever been made along the lines of classical work on natural ecosystems, such as the Cedar Bog Lake ecosystem of Minnesota (10) or the Silver Springs ecosystem of Florida (11).
Application of the energy budget concept
Complete or nearly complete energy budgets have been constructed for only six species of stored-grain insects. Of these, the pioneer study on the red flour beetle, Tribolium castaneum Herbst, was done by Klekowski and his group (12) in Poland, and the work on the remaining five species was done in the 1970s by our group in Winnipeg, Canada. The basic terminology of bioenergetics approved by the International Biological Program (13) and a brief account of the current state of knowledge of bioenergetics of stored-product insects are given below.
Several simple equations using the caloric unit per unit time for each animal or plant species are used to express an energy budget:
Material removed (MR) = unused material (NU) + consumption(C)
Consumption = production (P) + metabolism (R) + rejecta (FU)
Assimilation (A) = consuption - rejecta (FU)
Not all available food is used by a species of insect or plant. Potentially available food is either ignored or killed by an individual or a population (= trophic unit), thus removing it from a preceding trophic level (MR). A large part often remains unused (NU), and a small part is usually used (C).
Production means body growth including "skins" or silk shed by insects. Metabolism (or cost of maintenance of the species) includes respiration and physiological activity to maintain the body and its functions. Rejecta means that part of the ingested food that is eliminated as excement, such as urine and other excreted products. Assimilation is the sum of production and metabolism.
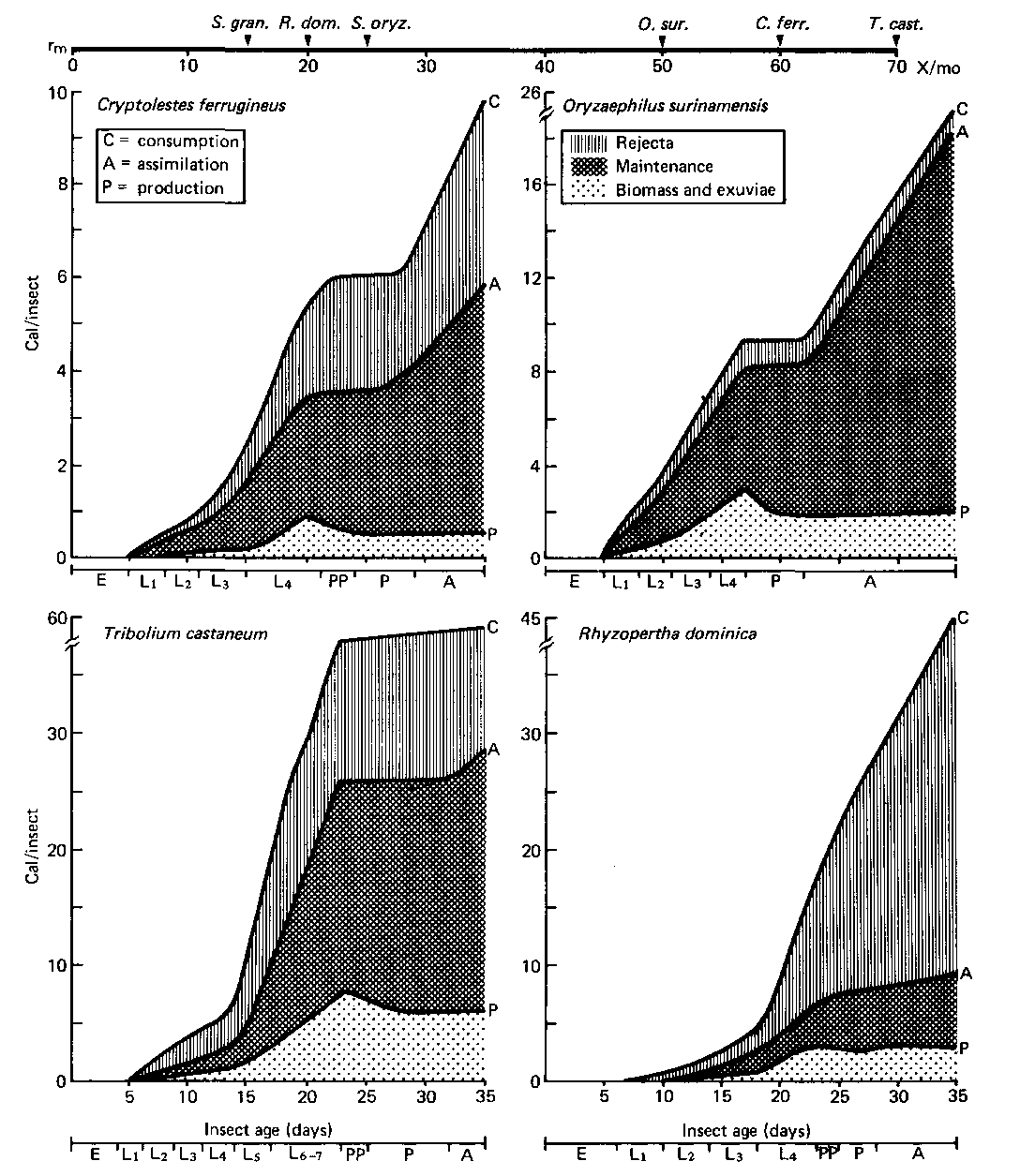
In the post-harvest stored-grain ecosystem, the primary producers of the first trophic level are crop seeds. Food energy is transferred from these basic energy units by pest organisms. The approximate caloric content per kernel for wheat is 124 cal, corn {maize) 564 cal, sorghum 104 cal, millet 35 cal. Most energy budget studies carried out so far have been done on wheat kernels. The insect energy budgets (figs. 2 and 3) are based on cumulative measurements of caloric values of each developmental age interval from egg to adult. Each insect was allowed to complete its life history while feeding on a single kernel or a measured quantity of cereal product. The general experimental procedure for these studies can be obtained from the original papers cited in the references
Briefly, construction of an energy budget involves: rearing the insect singly through all its life stages using 40-100 replications for each life stage; separating various come portents, such as uneaten food, faeces, exuviae, and the body, drying and then weighing them with a precision microbalance. The caloric values are determined with a microbomb calorimeter from combustion of pellets made out of dried material. Respiration of each age group is determined with a respirometer, such as a Warburg apparatus. The calculations are then made from these data.
Some experimental results
Food energy consumption of the larval stage ranged from a minimum of 5.8 cal per individual for Cryptolestes ferrugineus Stephens to a maximum of 78.7 cal per individual for Sitophilus granaries L.; the daily adult consumption is, however, highest for Rhyzopertha dominica F. (table 1) 114-17). The degree of consumption seems to be generally correlated with the body weights of the insects, which are 0.107 mg for C. ferrugineus and 1.226 mg for S. granaries adults. These figures represent the minimum and maximum of the range (table 1).
The number of calories used for respiration by immature specimens was the highest for S. granaries at 53.25 per individual, and lowest for C. ferrugineus at 3.82 per individual. An adult S. granaries also required the most energy, 1.2 cal per day; Tribolium castaneum Herbst was the second highest user of calories, and C. ferrugineus was the lowest user, 0.07 cal per day (table 2) (12, 14-17).
The mean biomass of various life stages of five species of stored-product beetles are compiled in table 3. In the egg stage, Oryzaephilus surinamensis L. had the highest biomass, 0.0126 mg; S. granaries had the highest biomass for all other life stages. Again, the size differences seem to be the reason for the differences in biomass among species.
The cumulative energy budgets of six species of stored. product insects are illustrated in figures 2 and 3. A comparison of these figures shows that, although S. granaries removed the maximum amount of gross food energy from cereal grains, O. surinamensis assimilated food energy most efficiently (fig. 2; little energy is released as rejecta or excrement). The most wasteful feeder was R. dominica because it assimilated a small part of what it voraciously ate (fig. 2; note the ratio between rejecta, including frass, and maintenance part of the diagram). Although Tribolium castaneum Herbst is the most adaptive (plasticity index = 700 [18]) and prolific (intrinsic rate of increase = 70 X per month [19]) stored-product insect species, Klekowski et al. (12) failed to measure its daily food intake (consumption), thereby reducing the usefulness of their otherwise excellent study.
TABLE 1. Mean Consumption of Food by Five Stored-Product Beetles in Immature and Adult Stages at 30 C
| Insect | Stage | Mean
adult weight (mg) |
Consumption (cal/ individual) |
Feeding period (days) |
Food | Source |
| Cryptolestes ferrugineus |
immature | 5.80 | 30 | whole wheat | ||
| adult | 0.107 | 0.59 | 1 | whole wheat | ref. 14 | |
| Oryzaephilus surinamensis |
immature | 9.17 | 22 | rolled oats | ||
| adult | 0.296 | 1.07 | 1 | rolled oats | ref.15 | |
| Rhyzopertha dominica |
immature | 21.00 | 30 | whole wheat | ||
| adult | 0.345 | 3.91 | 1 | whore wheat | ref. 14 | |
| Sitophilus | immature | 78.70 | 30 | whole wheat | ||
| granaries | adult | 1.226 | 3.55 | 1 | whore wheat | ref. 16 |
| Sitophilus | immature | 31.10 | 30 | whole wheat | ||
| oryzae | adult | 0.609 | 0.78 | 1 | whole wheat | ref. 17 |
TABLE 2. Respiration by Immature and Adult Specimens of Six Stored-Product Beetles at 29-30 C and 70 or 80 per cent RH
| Insect | Stage | Period (days) |
Food | Respiration (cal/individual) |
Source |
| Cryptolestes | immature | 30 | whole wheat | 3.82 | |
| ferrugineus | adult | 1 | whore wheat | 0.07 | ref.16 |
| Oryzaephilus | immature | 22 | rolled oats | 6.23 | |
| urinamensis | adult | 1 | rolled oats | 0.53 | ref. 15 |
| Tribolium | immature | 32 | wheat flour + brewers' Yeast | 21.0 | |
| Castaneum | adult | 1 | wheat flour+ brewers' yeast | 0.87 | ref.12 |
| Rhyzopertha | immature | 30 | whole wheat | 5.36 | |
| Dominica | adult | 1 | whore wheat | 0.35 | ref. 14 |
| Sitophilus | immature | 30 | whole wheat | 53.25 | |
| granarius | adult | 1 | whole wheat | 1.20 | ref.16 |
| Sitophilus | immature | 30 | whole wheat | 20.03 | |
| Oryzae | adult | 1 | whore wheat | 0.38 | ref.17 |
TABLE 3. Mean Biomass (Dry Weight, mg x 10-3) of Various Life Stages of Five Stored-Product Beetles
| Egg | L1 | L2 | L3 | L4 | PP | P | A | |
| Cryptolestes ferrugineus | 2.8 | 6.2 | 13.5 | 41.5 | 108.9 | 124.5 | 102.5 | 106.9 |
| Oryzaephilus surinamensis | 12.6 | 16.2 | 49.5 | 167.0 | 244.3 | - | 250.7 | 296.1 |
| Rhyzopertha dominica | 4.5 | 11.3 | 22.3 | 101.4 | 325.1 | 455.3 | 339.9 | 345.0 |
| Sitophilus granarius | 10.8 | 42.1 | 98.2 | 344.3 | 1305.5 | 1694.6 | 1482.2 | 1226.4 |
| Sitophilus oryzae | 6.3 | 16.9 | 48.9 | 137.7 | 632.3 | 961.2 | 656.7 | 609.0 |
L1 - L4. indicate first to fourth larval
instars; PP, prepupa; P. pupa; A, adult.
Sources: as in table 1.
The relative efficiency of food energy transfer can be best compared by ranking the six species of stored-product insects on the basis of two criteria - their net production efficiency (P/A x 100) and cumulative assimilation efficiency (A/C x 100)-as given in table 4 (12,20). A very active insect that is capable of generating heat readily in stored cereal and its products (21,22), O. surinamensis has the highest score on both of the efficiency criteria. Sitophilus oryzae, which is one of the most destructive stored-product insects in Africa, Southern Asia, and South
America (18), has the second highest assimilating efficiency by immature specimens (80 per cent), even though the net production efficiency of adults was only 12 per cent as compared to 47 per cent for O. surinamensis.
Campbell and Sinha (14) showed that, under similar environmental conditions, different species of stored-product beetles demonstrate a close relationship between energy conversion efficiency of assimilated food energy into egg production and population increase. When the daily net production for the first 10 to 20 days of adult life of five species (see list of species names in table 2 except O. surinamensis) was compared with their calculated intrinsic rate of population increase (table 5) (19,23-31), a linear relationship with R2 = 0.928 was found (regression calculated by the least-square method). From this relationship they postulated that the more efficient an adult beetle is in transforming assimilated energy into egg energy, the higher will be its rate of population increase. The predicted net production efficiency figure for adult O. surinamensis is about 32 per cent in relation to its intrinsic rate of increase (50 X per month, figure on Campbell and Sinhas' regression curve), falling below the predicted figures for T. castaneum and C. ferrugineus. The experimentally obtained value of 47 per cent indicates that future modification of the curve may be necessary and that it may be accurate only for species reared on identical food.
TABLE 4. Net Productivity Efficiency of Young Adults and Cumulative Assimilation Efficiency of Immature Stages of Stored-Product Beetles
| Net production efficiency of adults(%) | Cumulative assimilation efficiency of immature specimens (%) | Food used | |
| Cryptolestes ferrugineus | 22 | 72 | whole wheat |
| Oryzaephilus surinamensis | 47 | 90 | rolled oats |
| Rhyzopertha dominica | 15 | no determination | whole wheat |
| Tribolium castaneum | 45 | 46* | flour + brewers'yeast |
| Sitophilus granaries | 5 | 76 | whole wheat |
| Sitophilus oryzae | 12 | 80 | whole wheat |
* Consumption was estimated by Klekowsi et al.
(12) using data from Tenebrio molitor L. (20).
Sources: as in table 1.
TABLE 5. The Range of Temperatures, Minimum Humidity, and Rate of Increase in 28 Days for Six Stored-Product Beetles
| Insect | Temperature (°C) | Intrinsic rate of increase (rm) per lunar month | Source | ||
| Min. | Optimum | ||||
| Cryptolestes ferrugineus | 23 | 32-35 | 10 | 60 | refs. 23, 24 |
| Oryzaephilus surinamensis | 21 | 31-34 | 10 | 50 | refs. 25 |
| Tribolium castaneum | 22 | 32-35 | 1 | 70 | refs. 26-28 |
| Rhyzopertha dominica | 23 | 32-35 | 30 | 20 | refs 29 |
| Sitophilus granariues | 15 | 26-30 | 50 | 15 | refs. 30, 31 |
| Sitophilus oryzae | 17 | 27-31 | 60 | 25 | refs. 27, 29 |
Compiled by Howe (19).
Few generalizations can be made from our limited knowledge of the bioenergetics of the stored-product insects and the nature of the energy flow in the stored grain ecosystem of which they are a part. Unfortunately, bioenergetic studies done to date give energy flow data only at the level of individual insects. Until energy budgets at the population, community, and ecosystem levels involving insects, mites, and micro-organisms are done, extrapolations may be made only on the food energy consuming capacity of groups of beetles of a single species for a known quantity of grain stored in a granary; for example, the caloric energy used by 10,000 individuals of S. granarius in a tonne of wheat stored at 30 C.
In his scholarly analysis of Indian agro-ecosystems, Mitchell (4) cited the energy budget of the rice weevil, S. oryzae, prepared by Sigh et al. (17), as an excellent example of how to do an energy budget that could help others to see its applicator in both abstract ecology and in agriculture. He feels that this type of energy budget can be adapted to an agro-ecosystem model. When the number of beetles per unit of food resource is known, then the input can be specified as the number of beetles times the calories ingested per beetle. The output to decomposers will be the product of the numbers times the yield of excrete, and the energy in growth will eventually be transferred to consumers or decomposers.
In a world with a chronically short food supply, management strategy of stored food in the form of food energy conservation is likely to be more sophisticated in the future. Accurate prediction on a quantitative basis of potential loss in stored food in each climatic zone of the world may soon be possible with use of computer simulation models. Basic data on bioenergetics compiled in this report and those on intrinsic rate of natural increase of insects (19) and other storage pests will be essential to realize this goal.
In conclusion, it has now been demonstrated that the measurement of energy removed from grain kernels and their products by insect infestation can be quantified in caloric values. Pathways of the energy transferred from human and livestock food to the metabolic activities and body tissues of pest organisms can be accurately charted. This information can then be used in conjunction with other biological and ecological characteristics of the pest species (e.g., intrinsic rate of increase) and the stored food grain to develop a uniform strategy for assessing and reducing food loss.
References
1. D.W. Hall Handling and Storage of Food Grains in Tropical and Subtropical Areas FAO Agr. Dev. Paper No. 90 (FAO, Rome, 1970).
2. K. L. Harris and C.J. Lindblad, Postharvest Grain Assessment Methods (Amer. Assoc. of Cereal Chemists, St. Paul, Minn., USA, 1979).
3. E.P. Odum, Fundamentals of Ecology, 2nd ed. (W. B. Saunders Co., Philadelphia, Penn. USA, 1959).
4. R. Mitchell, The Analysis of Indian Agro-ecosystems (Inter-print, New Delhi, 1979).
5. R.N, Sinha, "Ecology of Microflora in Stored Grain," Ann. Technol. Agric., 28: 191-209 (1979).
6. R. Margalef, "On Certain Unifying Principles in Ecology" Amer. Nat., 97: 357-374 (1963).
7. R.N. Sinha, H.A.H. Wallace, and F. Chebib, "Principal-Component Analysis of Interrelations among Fungi, Mites, and Insects in Grain Bulk Ecosystems," Ecology, 50: 536-547 (1969).
8. J.E.M.H. Bronswijk van and R.N. Sinha, "Interrelations among Physical, Biological, and Chemical Variates in Stored-Grein Ecosystems - a Descriptive and Multivariate Study," Ann. Entomol. Soc. Amer., 64: 789-803 (1971).
9. N.D.G. White and R.N. Sinha, "Changes in Stored Wheat Ecosystems Infested with Two Combinations of insects Species," Can. J. Zool., 58: 1524-1534 (1980).
10. R.L. Lindeman, "The Trophic-Dynamic Aspect of Ecology," Ecology, 23: 399-418 (1942).
11. H.T, Odum, "Trophic Structure and Productivity of Silver Springs, Florida," Ecol. Mono., 27: 55-112 (1957).
12. R.Z. Klekowski, T. Prus, and H. Zyromska-Rudzka, "Elements of Energy Budget of Tribolium castanoum (Hbst.) in Its Developmental Cycle," in K. Petrusewicz, ea., Secondary Productivity of Terrestrial Ecosystem, (PWN, Warsaw, Krakow, 1967), pp. 859-879.
13. K. Petrusewicz and A. Macfadyen Productivity of Terrestrial Animals, Principles and Methods IBP Handbook No. 13. (Blackwell Publ, Oxford, 1970).
14. A. Campbell and R.N. Sinha, "Bioenergetics of Granivorous Beetles, Cryptolestes ferrugineus and Rhyzopertha dominica (Coleoptera: Cuculidae and Bostrichidae)," Can. J. Zool., 56: 624-633. (1978).
15. N.D.G. White and R.N. Sinha, "An Energy Budget for Oryzaeophilus surinamensis (Coleoptera: Cuculidae) Feeding on Roiled Oats," Environ. Entomol. (in press).
16. A. Campbell, N.B. Singh, and R,N. Sinha, "Bioenergetics of the Granary Weevil, Sitophilus granarius (L.) (Coleoptera: Curculionidae), Can. J. Zool., 54: 786-798 (1976).
17. N.B. Singh, A. Campbell, and R.N. Sinha, "An Energy Budget of Sitophilus oryzae (Coleoptera: Curculionidae)," Ann. Entomol. Soc. Amer., 69: 503-512 (1976).
18. R.N. Sinha, "Climate and the Infestation of Stored Cereals by Insects," Proc. 1st Int. Working Conf. Stored Prod. EntomoL, Savannah, Georgia (1974), W. 117-141.
19. R.W. Howe, "A Summary of Estimates of Optimal and Minimal Conditions for Population Increase of Some Stored Products Insects. J. Stored Prod. Res., 1: 177-184 (1965).
20. A.C. Evans and E.R. Goodliffe, "The Utilization of Food by the Larva of the Mealworm Tenebrio molitor L," Proc. R. Entomol. Soc. London, Ser. A, 14: 57-62 (1939).
21. M.T. Armstrong and R.W. Howe, "The Saw-Toothed Grain Beetle Oryzaephilus surinamensis in Home-Grown Grain," J. Agr. Eng. Res., 8: 256-261 (1963).
22. R.N. Sinha and H.A.H. Wallace, "Ecology of Insect-lnduced Hot Spots in Stored Grain in Western Canada," Res. Popul. Ecol., 8: 107-132 (1966).
23. G.W. Bishop, "The Comparative Bionomics of American Cryptolestes (Coleoptera, Cuculidae) that Infest Stored Grain," Ann. Entomol. Soc. Amer., 52: 657-665 (1959).
24. L.B. Smith, "The Intrinsic Rate of Natural Increase of Cryptolestes ferrugineus [Stephens) (Coleoptera: Cuculidae)," J. Scored Prod. Res., 1: 3549 (1965).
25. R.W. Howe, "The Biology of the Two Common Storage Species of Oryzaphilus (Coleoptera, Cuculidae)," Ann. Appl. Biol., 44: 341-355 (1956).
26. R.W. Howe, "The Effect of Temperature and Humidity on the Rate of Development and Mortality of Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae)," Ann. Appl. Biol., 44: 356-368 (1956).
27. R.W. Howe, "The Effects of Temperature end Humidity on the Oviposition Rate of Tribolium castaneum (Hbst.) (Coleoptera Tenebrionidae)," Bull. Entomol. Res., 53: 301310 (1962).
28. R.H. Leslie and T. Park, "The Intrinsic Rate of Natural Increase of Tribolium castaneum Herbst," EcoIogv, 30: 469477 (1949).
29. L.C. Birch, "Experimental Background to the Study of the Distribution and Abundance of Insects - I. The Influence of Temperature, Moisture and Food on the Innate Capacity for Increase of Three Grain Beetles," Ecology, 34: 698-711 (1953).
30. L.E.S. Eastham end S.B. McCully, "The Oviposition Responses of Calandra granaria Linn."J. Exp Biol., 20: 35-42 (1943).
31. L.E.S. Eastham and F. Seagrove, "The Influence of Temperature and Humidity on Instar Length in Calandra granaria Linn," J. Exp. Biol., 24: 79-94 (1947).
{kind=link}
{kind=link}
{kind=link}