Premature infants,
presently better defined as preterm infants born before 38 weeks
of gestation, present an interesting model to assess effects of
protein and energy intakes on growth. Preterm infants are
affected by significant illness usually associated with
immaturity. They often become malnourished because of
intercurrent morbidity or because of inappropriate management.
Such infants, when fed adequately, grow at a fast rate, but
usually do not fully catch up (HACK et al., 1984;
GEORGIEFF, ZEMPEL and CHANG, 1989).
Advances in neonatal clinical practices have allowed for the survival of progressively smaller infants. The overall survival rate for very low birth weight (VLBW) infants (birth weight <1500 g) is 85-90%, and for those < 800 g it is close to 60% (HACK et al., 1991b). For VLBW infants the initial days or weeks of life are characterized by significant weight loss due to catabolic illness and insufficient protein-energy supply. VLBW infants constitute in most industrialized countries 1.01.5% of all births and account for a significant proportion of neonatal and infant mortality. The accepted practice of modern neonatal care allows for a 12-15% body weight loss over the initial 10 days. After parenteral and enteral nutritional support is given, they will take 15-20 days to regain birth weight. The catch-up growth observed in these infants may last 3-8 weeks. During this time they will grow at rates of 15-30 g/kg/d, which is double the normal growth rate in utero and three times that observed in the full-term newborn. Nutritional practices have had a significant impact in shortening duration of hospitalization and improving survival.
The criteria for discharge include not only attaining basic physiologic maturity but also reaching a critical weight, usually 1800-2200 g, depending on individual center practices. Thus, growth rate for VLBW infants has medical as well as economical implications. The duration of nutritional deprivation and time to recover linear and head growth has been correlated with subsequent developmental outcome (HACK et al., 1991a; The Infant Health and Development Program, 1990). It is for these reasons that the study of nutrient needs for optimal catch-up growth of these infants has been the focus of extensive research efforts over the past decades.
The results of multiple studies provide a wide range of recommended energy and protein intakes; they vary from 110-150 kcal/kg and from 2.5-4.2 g of protein/kg daily (FOMON and HEIRD, 1986). The main sources of variance in these studies are related to fecal losses and the allowance for growth. The resting metabolic rate, including post-prandial thermogenesis, in most studies is 50-60 kcal per kg. The expenditure related to minor thermal stress despite a controlled environment is 10 kcal per kg. An additional 10 kcal/kg are provided for activity, fecal losses vary from 10 to 40 kcal/kg depending on what is fed, and the allowance for synthesis and storage (growth) is 35-60 kcal/kg. Most studies have varied both the amount of calories and protein fed, and it becomes extremely difficult to evaluate the effect of P/E ratios on weight gain and catch-up growth (FOMON and HEIRD, 1986).
Another key problem in evaluating these data is establishing the goal(s) of nutritional recovery in these infants. The present goal proposed by most national and international expert committees is a logical one: VLBW and LBW infants should attain as early as possible a body weight approximating that of a normal fetus of the same conceptional age (AAP, 1985; ZIEGLER, 1986; HEIRD and KASHYAP, 1989). A second goal that is often not specified is that the quality of the tissue gain be similar to that accreted normally in utero during the equivalent gestation period (the ratio of lean to fat stored at 32 weeks is approximately 1/1). The body of available information indicates that the first goal is achievable with presently available formulations (BROOKE, ALVEAR and ARNOLD, 1979; REICHMAN et al., 1981; REICHMAN et al., 1982; CHESSEX et al., 1983; PUTET et al., 1984).
These studies also show that in most cases the fat/lean ratio of tissue gained ex utero is higher than what is normal in utero. The higher fat/lean ratio of weight gain of VLBW infants during catch-up can be interpreted as an unavoidable consequence of post-natal nutrition and growth regulation. Alternatively, it may be interpreted as resulting from inadequate nutrient supply for optimal lean tissue accretion. Heird and others have proposed that this problem results from the absolute energy supplied and the P/E ratio of the formula, while others have suggested that micronutrients may be critical to optimize lean tissue gained.
Recent experimental observations in VLBW infants indicate that it is feasible, by a gestational age of 40 weeks ex utero, not only to attain but even exceed growth rates in utero while at the same time adding sufficient lean tissue to reach a weight and body composition that are the equivalent to those of a normal baby at birth (PUTET et al., 1984; KASHYAP et al., 1986; 1987; 1988). This required not only a higher energy intake (150 kcal/kg rather than 98 kcal/d) but also that protein be increased from an intake of 3.3 g/kg to an intake of 4.5 g/kg/d. The authors conclude that a high energy intake is required to optimize protein utilization for optimal catch-up. This study is one of the few that varied energy and protein intakes independently.
The observations also show a significant correlation between the urinary excretion of C-peptide and weight gain. The high-energy, high-protein group had significantly higher C-peptide excretion, suggesting that insulin response was greater. The accretion of fat was higher than in utero, yet since the normal fetus will increase fat gain during the last weeks of gestation, it is possible that with the high-protein, high-energy diets the relative excess of fat will decrease with advancing age.
Based on metabolic balance and growth observations on over 100 infants, Heird has proposed and validated simple linear equations that relate weight gain of VLBW infants during recovery to energy intake, protein intake and birth weight; similarly, N retention can be predicted from intake, and energy stored predicted from energy intake. These equations permit the estimation of lean tissue and fat tissue gained from energy balance and N-balance data, which in turn are predicted from N and energy intake.
A prospective experimental validation of this approach to meet the goals of optimal nutrition for VLBW infants has recently been reported (HEIRD and KASHYAP, 1989). The observations demonstrated that, on high-energy (142 kcal/kg) and high-protein (4.2 g per kg) diets, full catch-up is possible, and lean tissue accretion paralleling in utero rates is feasible. The only undesirable effect, as predicted from the equations, is excessive fat storage. In attemps to improve catch-up growth of VLBW infants, a pending issue is how to enhance protein utilization without increasing energy stored as fat.
The use of high-carbohydrate, low-fat energy sources can be proposed as the next logical step based on the known effects of carbohydrates on insulin and IGF release (BRESSON et al., 1989). Another approach might be to intervene by adding factors that enhance N retention; growth hormone, IGF-I and insulin would be likely candidates (ARIZTIA et al., 1969; MONCKEBERG et al., 1963; PREVOST-THIERIOT, 1988). The use of specific amino acid blends that induce anabolic hormones might be yet a third approach to attain this goal; arginine and the branched-chain amino acids may be considered, since under catabolic conditions they may promote N retention.
The use of high-carbohydrate diets may pose special problems for infants with cardiorespiratory problems, since CHO fuel oxidation leads to higher carbon dioxide formation and may also increase energy expenditure, if the excess energy is converted to fat rather than contributing to lean tissue synthesis (KURZNER et al., 1988). This is an area that clearly needs further investigation. The results may offer new insights on how to optimize lean body mass accretion in malnourished infants and children with primary and secondary malnutrition.
The use of whole-body protein and nitrogen kinetic techniques has led to a clearer understanding of P/E relationships in the neonate as well as in other groups. Preterm infants have been preferred experimental subjects for these studies since they are small and have high turnover rates. Thus, they require less isotope dose and will attain steady state faster than older subjects. They are usually on controlled intakes, and since less is known of their true requirements it is ethically permissible to evaluate ranges of intakes that may have an impact on metabolic outcomes.
The published information indicates that the low birth weight infant has an extraordinarily high rate of protein synthesis. When 13C plasma leucine is used to assess synthesis, the reported rate is 5-10 g/kg/d, whereas using 15N glycine and urinary urea enrichment the values have ranged from 13 to 26 g of protein/kg/d (NICHOLSON et al., 1970; BIER and YOUNG, 1986; DENNE and KALHAN, 1987). The variance in these studies can be explained by clinical and experimental conditions under which the studies were conducted.
In any case, the observations indicate that both synthesis and breakdown are extraordinarily active and that only a small proportion of what is synthesized is actually stored. If 12 g are synthesized and 2 g stored then 10 g must be catabolized; thus, only about 20% of synthesis is actual storage. Protein synthesis is an energy-demanding process, and the energy cost of tissue gain in the neonate must include this cost. WATERLOW (1986) has suggested that close to half of the cost of lean body mass gain in the neonate can be explained by the energy cost of protein synthesis.
This has been validated by experimental studies demonstrating that for each 1 g of protein stored, 5 g are synthesized. This resulted in an energy cost of 10 kcal, and thus 2 kcal were required per g of protein synthesis (CATZEFLIS et al., 1985; BEAUFRERE et al., 1990). This may be considered wasteful but, on the other hand, the system, as it operates, offers multiple sites for regulation and adaptation that would otherwise not be available.
The high protein turnover rate favors tissue remodeling and appears to be a precondition for rapid growth. In the final analysis, it is the balance between synthesis and catabolism which determines protein accretion. Dietary energy, protein, and hormonal regulators affected by maturation and nutrient supply affect rates of synthesis and breakdown. Two mechanisms may account for net protein accretion in response to diet: either protein synthesis increases or protein breakdown decreases. Of course, both may also occur concomitantly.
The evidence available from leucine oxidation kinetics indicates that in the full-term infant it is the activation of synthesis after feeding which determines protein accretion, but in the premature infant, where synthetic rates are extremely high, it is the inhibition of breakdown after feeding which determines net accretion (DENNE, KARN and LIECHTY, 1991a; c). It has been suggested that, if protein synthesis is low, accretion after feeding will be mediated by increases in synthetic rates; conversely, if synthetic rates are already maximal, the inhibition of breakdown will serve to increase accretion.
A few studies in the newborn have addressed the effect of protein-energy interactions on turnover rates. The available information from these studies indicates that energy supply over 60 kcal/kg is needed to sustain protein synthesis, and that amino acid quantity, quality and route of delivery affect turnover rates (DENNE, KARN and LIECHTY, 1991b). A controlled study demonstrated that rates of synthesis and breakdown as measured by 15N glycine were not affected in premature infants receiving 60 vs 90 kcal/kg/d parenterally (DUFFY et al., 1981). Less is known on the effect of the type of non-protein energy sources on turnover rates, although the effect of fuel sources on energy balance has been verified in the neonate (DENNE, KARN and LIECHTY, 1991a). A recent report suggests that fuel source affects amino acid oxidation rates and protein breakdown but not synthetic rates. Infants given parenteral glucose and lipid had lower rates of breakdown relative to a group receiving only glucose as a source of non-protein energy (BRESSON et al., 1991).
The special
case of the small-for-gestational-age (SGA) LBW infant has also
been evaluated, demonstrating that synthesis and breakdown rates
are lower than for appropriate-for-gestational-age (AGA) infants
(CAUDERAY et al., 1987). The SGA (birth weight 1500 g;
gestational age 35 wks) group demonstrated lower synthetic rates
and a 20% higher protein storage per gram of protein synthesis
relative to weight matched AGA (gestational age 32 wks) infants
(MICHELI and SCHUTZ, 1987; MICHELI et al., 1990). This
lower synthetic rate needs to be interpreted on the basis of the
known decrease in synthetic rates with advancing maturation.
Thus, the lower rate may be similar to that of infants of
equivalent maturation. The experimental data show a close
relationship between protein synthesis, protein accretion, and
energy expenditure suggesting that these processes are intimately
linked.
Virtually all children with
severe protein-energy malnutrition (PEM) have deficits in both
total body fat and total body protein. Nutritional rehabilitation
aims to replace these deficits as rapidly as possible.
Historically, the emphasis in treatment was placed on high
protein intakes, but it was later recognized that calories and
not protein were often the limiting factor in the rate of weight
gain during recovery (DEAN and SKINNER, 1957). The efficacy of
high-energy feeding to achieve rapid weight gain during recovery
from PEM is well documented (WATERLOW, 1961; JACKSON and WOOTTON,
1990). What is less certain is the relationship between the high
energy intake and the composition of the tissue being laid down
during rapid catch-up growth in the child recovering from PEM.
The increase in body weight during recovery from PEM may be regarded as equal to the increment in lean tissue plus the increment in fat tissue. The former consists of the increase in muscle mass plus the increase in non-muscle lean tissue. This latter increase is small compared to the increase in muscle mass, and the weight gained during recovery may be equated to the increase in fat tissue plus the increase in muscle tissue (JACKSON et al., 1977).
Fat has an energy content of 9.4 kcal/g and protein 5.7 kcal/g. Assuming that fat tissue has 15% as water, the stored energy of tissue fat is 8 kcal/g. Similarly, since lean tissue has an average protein content of 20%, it has an energy storage value of 1.2 kcal/g. A hypothetical line can be constructed using the axes, cost of growth, and increase in muscle mass expressed as a percentage of the increase in body weight.
Thus, if all the tissue laid down were fat tissue, and none of the increase in body weight were due to new muscle tissue, the cost of growth would be 8 kcal/g. Conversely, if all the new tissue gained were lean, the cost of growth would be 1.2 kcal/g (JACKSON et al., ROBERTS and YOUNG, 1988).
Muscle tissue is reduced by up to 70% in children with severe PEM; it provides the largest store of potentially available protein in the body. During recovery from malnutrition there is a significant: increase in muscle mass. This increase is proportionately greater than the increase in body weight and may differ according to the initial degree of stunting or wasting (REEDS et al., 1978). The evidence suggests that both hypertrophy and hyperplasia must take place, but the extent to which each process contributes to the muscle repletion has not been clarified.
Malnourished infants during the period of rapid catch-up growth are able to accept higher energy intakes when fed high energy density diets (ASHWORTH, 1976). It is not clear how the effect of the energy density of the feed affects the control of intake, nor is it clear if this effect is independent of the protein content of the diet. There is good evidence of spontaneous regulation of energy intake from the first week of life in full-term infants (FOMON, 1967). Both AGA and SGA infants consume a greater volume of a low energy density regimen than high energy density regimen. During the first 4 to 6 weeks of life, the reduction in intake after a high-energy feed is not sufficient to compensate fully for the difference in energy content; thus, higher energy intake results when a high energy density formula is given than when a low-energy feed is provided. After 6 to 8 weeks, normal infants are able to compensate, and there is no difference in energy intakes on feeds of different energy densities. We have observed that while most malnourished infants decrease their energy intake after normal weight-for-length is achieved, some maintain a high energy intake beyond the repletion phase. That is, they exceed normal weight-for-length by increasing adipose tissue mass (CHAVEZ et al., 1980). Thus, the set point for appetite regulation during recovery from malnutrition may be affected by conditions related to the infant or to the diet (Figures 1 and 2).
Energy and protein metabolism are closely interrelated (MUNRO, 1964). Several short-term studies have shown that the protein intake required for nitrogen equilibrium decreases with increasing energy intake. This occurs not only when the initial energy intake level is deficient but also when intake exceeds requirements. On the other hand, a reduction of protein intake while energy intake remains constant can reduce the rate of growth in terms of weight and height, even when nitrogen balance is not negative. The mechanisms producing these effects are not clear. It is also known that with an energy intake that is borderline or inadequate, an increase in protein intake can result in improved weight gain. Energy requirements for maintenance of weight and normal body composition may be less when dietary protein is adequate. Except for post-prandial thermogenesis, protein intake does not have a specific effect on energy requirements (BROOKE and ALVEAR, 1982).
Anthropometry is expressed as mean values % of WHO median standard. After 2 months of age, catch-up weight was mainly fat, as evidenced by weight-for-length and triceps fat-fold gains (CHAVEZ et al., 1980).
Amino acid intake is the primary determinant of nitrogen retention when energy intake is fixed and adequate; conversely, an increased amino acid supply will be useless if energy is limiting (WATERLOW, 1986). Rapid rates of growth and of protein turnover require a generous energy supply. The energy cost of growth in infants with average composition of the tissue laid down has been estimated to be 5 kcal/g (FAO/WHO/UNU, 1985), of which about half can be accounted for by the energy content of the fat and protein that are deposited.
The requirements for catch-up growth can be estimated based on the available evidence. First, relatively more protein is needed for weight gain (assuming normal, balanced tissue gain) than for maintenance; therefore the P/E ratio in the diet needs to be higher than normal. Second, catch-up growth requires an increased intake of both energy and protein but the increase is several-fold greater for protein than for energy (WATERLOW, 1961).
The fact that undernutrition reduces the rate of growth must have been known for thousands of years, but malnutrition does more than this. By limiting the supply of nutrients for growth, it accentuates the development of those tissues which, at the time in question, have greater structural and perhaps metabolic stability than the rest (McCANCE, 1962). WIDDOWSON and McCANCE (1975) have described that there is a critical point in development when the size of an animal determines its appetite thereafter, and hence its rate of growth and dimensions at maturity. A small size at this critical time brought about by undernutrition is not followed by catch-up growth, however liberal the diet. A full diet produces catch-up growth only if the undernutrition, whatever its cause, has occurred after this critical period is over. Feeding can only restore a young animal to its percentile channel of growth and its ability to do this after long periods of undernutrition becomes progressively limited by the animal's chronological age. There is a time after which catch-up growth becomes impossible (WIDDOWSON and McCANCE, 1975).
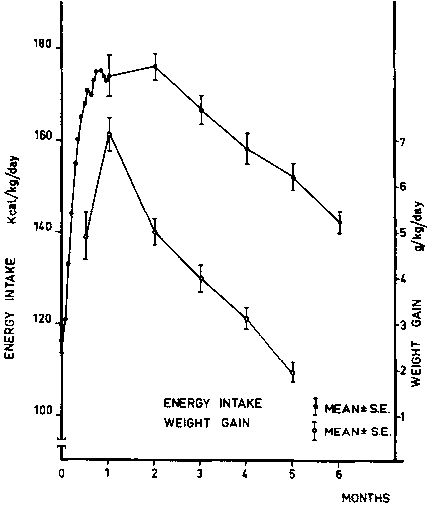
After 2 months of age, energy intake (solid circles) decreased as weight gain (open circles) slowed down (CHAVEZ et al., 1980).
ASHWORTH (1969) has described catch-up growth in malnourished infants of mean age 16.5 months, whose growth rates were fifteen times faster than those of normal children of similar ages and five times as fast as that of normal children of similar height or weight. Rapid growth was associated with high food intake (165 kcal and 3.8 g protein/kg body weight/d). This phenomenon occurred while infants were depleted and stopped upon reaching their expected weight-for-height (ASHWORTH, 1969). Children fed ad libitum a diet with 17% protein calories are able to regulate energy intake according to the demands of their body mass. If the P/E: ratio is lowered to 11%, they are able to consume 20% extra energy in an attempt to meet the level of protein necessary to reach maximal growth capacity determined by weight deficit (Rio et al., 1979).

Infants with high (> than median) energy intakes (open circles) during recovery had similar gain in weight-for-length than those with low (£ than median) energy intakes during the first 2 months. At this time the group with high energy intake exhibited gain-for-weight above the expected for given length. Intake data for the group as a whole corresponds to solid circles in Figure 3 (CHAVEZ et al., 1980).
Long-term observations of children severely malnourished in early life suggested permanent reductions in body length (STOCK and SMYTHE, 1967; GRAHAM, 1968). Other observations indicate that, if these children have the benefit of a dramatic improvement in their environment, they are able to exhibit catch-up growth (GRAHAM and ADRIANZEN, 1972; SCHUMACHER, PAWSON and KRETCHMER, 1987). In general, younger children tended to gain more length during recovery. This suggests that there is an influence of biological age on the capacity for catch-up growth (WALKER and GOLDEN, 1988).
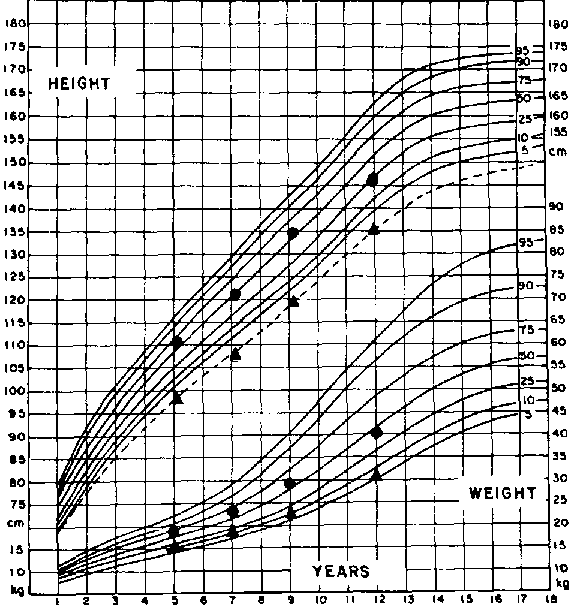
We have demonstrated that 180-220 kcal and 3.5 g protein/kg/d support a weight gain of 10-15 g/kg/d in infants under 2 years of age being treated for PEM. These infants have been followed for up to 15 years after discharge. A subgroup of infants gained weight beyond the median expected weight-for-length by the time of discharge, namely 110% of the WHO standard (Figure 3). On the first year follow-up, this group showed faster linear growth velocity than infants discharged with normal weight-for-length, but subsequently their growth velocity slowed down. Long-term follow-up of this cohort through adolescence demonstrated that they were shorter at 14 years of age than the group discharged with normal weight-for-length relationship (ALVEAR, VIAL and ARTAZA, 1991) (Figure 4). We have also described a relationship between arm fat area, length-for-age, bone age and weight-for-age suggesting that the extra amount of fat gained during early treatment accelerated bone age maturation. Thus, although faster growth was achieved in the short term, stunting in the long term was worse (ALVEAR et al., 1986).
The energy cost of repleting tissue deficit ranges from 4 to 5 kcal/g weight gain (KERR et al., 1973). The impressive gains in weight made by recovering malnourished infants are largely fat, and reconstruction of lean body mass does not occur equally at all rates of weight gain. Protein status may be maintained if protein in the diet is quantitatively and qualitatively sufficient.
If protein
intake is low or of poor quality, the development of obesity may
be accompanied by a simultaneous deterioration of lean body
composition (MacLEAN and GRAHAM, 1980). If protein intake is
increased to 4-5 g/kg/d and energy raised to 170 kcal/kg/d, it is
possible to obtain a weight gain of 11.8 g/kg/d, shortening the
time of treatment without increasing the fat percent of total
weight gained (FJELD, SCHOELLER and BROWN, 1989a). These authors
have also reported, using doubly-labelled water to assess energy
expenditure, that metabolizable energy needs for growth during
recovery from PEM can be predicted based on initial metabolic
mass, rate of weight gain, and composition of the weight gain
(FJELD, SCHOELLER and BROWN, 1989b). Several studies in animals
and humans including infants suggest that, on an isoenergetic
basis, carbohydrate is more effective than fat in supporting
protein retention (NOSE et al., 1987). Recent studies
suggest that N urea utilization in malnourished children is also
enhanced by carbohydrate feeding (JACKSON, 1989).
Protein intake is a major
determinant of growth in early life under health and disease
conditions. Adult patients with chronic renal insufficiency (CRI)
may benefit from moderate protein restriction by a decrease in
the rate of progression of renal insufficiency (BRENNER et
al., 1982). Recent studies in adults suggest that protein
intakes 30 to 50% below the RDA are associated with a significant
amelioration in the rate of progression of CRI (ROSMAN, 1984).
Studies in experimental animals support this contention, yet the
few studies performed in growing animals indicate that severe
protein restriction has an adverse impact on growth and does not
convey additional benefits in slowing progression of CRI beyond
those seen using moderate protein restriction.
Dietary protein intake in infants with CRI should serve to sustain normal growth and development, and at the same time preserve remaining renal function over the longest time possible. These two potentially conflicting objectives need consideration in defining an adequate protein intake for children with CRI (JUREIDINI et al., 1990). No specific protein intake recommendations are available for them. Most authors suggest that the existing RDAs for normal children be used, since they are considered safe (HELLERSTEIN, 1987). However, customary protein intakes of 'healthy' children after 6 months of life in the USA greatly exceed the existing national and international recommendations for protein intake.
We have recently completed a study to evaluate if a dietary protein intake given at levels recommended for 'healthy' children (P/E 5.6%) would sustain growth of children with chronic renal insufficiency, as compared to an intake close to that customarily consumed (P/E 10.4%) (UAUY et al., 1992). The primary outcome variable for growth evaluation was linear growth as measured by length-for-age and length gain velocity.
The initial 2 months prior to randomization (baseline period) were considered necessary for clinical stabilization and to standardize treatment including dietary management. All subjects received dietary protein corresponding to 8% of total energy. At 8 months of age, and for the next 10 months, the patients were randomized either to a low-dietary-protein group (P/E 5.6%) receiving an intake similar to that recommended by the USA NAS/NRC and international bodies (WHO/FAO/UNU), or to a control group (P/E 10.4%) receiving an intake close to that customary for American young children. These diets when consumed at 100 kcal/kg/d provided 1.4 and 2.6 g of protein per kg/d.
The calorie prescription for patients in the study was based on the RDA for normal infants of comparable length (RDA for length) rather than on actual weight of the subjects. The recommendation was that children consume 100-120% of the RDA for length. Protein intake of the study groups based on the formula composition and solid foods dropped from close to 2.0 to 1.4 g/kg/d in the low group and increased from 2.0 to 2.4 g/kg/d in the control diet group. The distribution around this mean indicates that during some months children in the low-protein group received as little as 1.0 or up to 1.8 g/kg/d. In the control group, the intakes ranged from 1.7 to 3.0 g/kg/d.
Mean energy intakes expressed per kg body weight oscillated around 100 kcal/kg/d, but expressed as % RDA for length the intakes were close to 90% of that considered ideal for 'healthy' children of equivalent length. Absolute weight, length, and head circumference increased significantly with time in both groups in a similar fashion. Mean weight gain after randomization to 18 months was 2.11 kg with the low-protein and 2.41 kg in the control-protein intake; the difference between diet groups was not significant. Length gain was 8.9 cm in the low-protein and 10.7 cm in the control group.
The repeated measure ANOVA indicated a significant time/diet interaction (p<0.083). The 1.8 cm difference in length gain between groups over the 10-months' diet intervention period was further explored using length gain velocity comparisons. The effects observed with absolute length were also evident in standardized length-for-age according to gender. The change in length SD score for age 8-18 months was -0.52 SD in the low and -0.04 SD in the control, for the 12-18 months' period the change was -0.06 in the low while it was +0.28 in the control diet group; this interaction of diet group and time was significant by repeated measure ANOVA. Length gain velocity over 6-months' periods was also expressed as SD score. The 0-6 months' data were analyzed based on the reported length at birth, while the 6-12 and 12-18 months' data were measured prospectively as part of the study (Figure 5). The interaction of diet, group and time was significant (p<0.04). Further analysis of this relationship was done using as covariates baseline weight, length, energy intake (expressed as kcal/kg/d and as % RDA for length) and change in length from birth. Baseline weight served to strengthen the effect of protein energy within this interaction model; the effect of diet group had a p < 0.036.
The remaining anthropometric measures showed time-related increases which were similar for both groups. The weight SD scores changed significantly over time for both, regardless of diet-protein level. For weight-for-length there was a significant decrease over time in both groups (p<0.016); the significance was given by the decrease observed from 6 to 8 months, that is, prior to being randomized to the dietary protein groups.
The results of this prospective randomized controlled trial of the effect of protein intake on growth and renal function of children with CRI clearly indicate that low protein energy diets within the range considered safe for 'healthy' children may in fact compromise linear growth of children with renal disease. This observation should be considered in light of the confounding effect of energy intake present in this study; namely, the difficulties in achieving the prescribed energy intakes may have determined a lower protein utilization than otherwise would be obtained if energy intake were sufficient.
As observed in this study, the compromise of linear growth in malnutrition is slow but pervasive. The end result is stunting. The problem relates not only to a few lost centimeters in length but, as suggested by multiple studies in malnourished children, poor linear growth due to lack of food or to chronic illness is subsequently associated with poorer intellectual function and lower capacity for physical work. The presently recommended intakes for normal children do not support catch-up growth of children with CRI. The results of this study lend further support to the use of length velocity SD score as a more sensitive indicator of growth failure and of potential nutritional inadequacies. The advantages of a low-protein diet for children with CRI are not clearly established, based on the available evidence. A moderately low dietary P/E ratio as demonstrated in this study may compromise linear growth without other evidence of protein deficiency.
{kind=link}
{kind=link}