This is the old United Nations University website. Visit the new site at http://unu.edu
Effects of vitamin-A status on hamster tracheal epithellum in viva in vitro
Luigi M. De Luca and Elizabeth M. McDowell
This paper highlights what is known about how vitamin A and the retinoids control epithelial morphology and function. The system of choice is the tracheal epithelium of the Syrian golden hamster. This species was originally selected by Saffiotti et al. [1] in their studies of chemical carcinogenesis of the respiratory tract because it is relatively resistant to respiratory infection. The system was then rendered more amenable to in vitro investigation by Sporn et al. [2], who defined conditions for maintaining the trachea in organ explant culture, as well as the tissue's requirement for retinoic acid (RA) in the maintenance of normal mucociliary differentiation. Following in viva observations [3; 4], McDowell et al. have recently used epithelial cell culture techniques to define the mucous cell as the target of vitamin A [5].
In vivo effect of vitamin-A status on the hamster tracheal epithelium
The final effect of nutritional deficiency of vitamin A on the tracheal epithelium is the replacement of ciliated cells and normal mucous cells (fig. 1) by squamoid cells (altered mucous cells), which normally characterize the epidermoid type of differentiation. Figure 2 shows the extent of such replacement, which results in the near occlusion of the tracheal lumen. Accumulation of bacteria and other external material influences survival of the animal, which eventually succumbs to infection.
The terminal stage is preceded by primary effects of the deficient diet. Careful monitoring of the changes in body weight and serum retinal levels allowed the definition of a stage of "minimal morphological change," which preceded the loss of body weight [3; 4; 6]. Measurement of the cell-division rates in the mucous and basal cells during the development of vitamin-A deficiency revealed that the rates (mitotic rates) were lowered in a non-uniform manner. The replication of mucous cells was profoundly reduced compared with that of the basal cells during the stage of "minimal morphological change." This is clearly shown in table 1.
This work leads to the conclusion that vitamin-A deficiency depresses epithelial cell division before epidermoid metaplasia formation is evident. If vitamin-A deficiency continues, however, the altered mucous cells regain the capacity to divide and the epithelium is replaced by flat "squamoid cells." These cells generally arise as a consequence of cell injury, whether caused by carcinogen exposure, mechanical injury, or, as in this case, nutritional deficiency (fig. 3). It is clear, then, that squamoid cells (but not columnar mucous cells) can survive and multiply in the absence of vitamin A. Reversal of squamous metaplasia to the normal mucociliary phenotype of the tracheal epithelium is only possible in the presence of vitamin A or one of its biologically active analogues.
TABLE 1. Proportions and mitotic rates of tracheal basal, mucous, and ciliated cells of control and vitamin-A-deprived hamsters
| Cell type | Control | Vitamin-A-deprived | P value |
| Proportion of total (%) | |||
| Basal | 28.7 ± 2.5 | 39.7 ± 2.7 | <.0001 |
| Mucous | 59.3 ± 2.6 | 53.1 ± 1.9 | <.001 |
| Ciliated | 11.0 ± 3.1 | 6.8 ± 1.8 | <.05 |
| Mitotic rate (% of total)a | |||
| Basal | 0 61 ± 0.13 | 0.29 ± 0.35 | <.062 |
| Mucous | 2.42 ± 1.19 | 0.15 ± 0.10 | <.01 |
| Ciliatedb | 0 | 0 | |
| Mitotic rate (% of own cell type)c | |||
| Basal | 2.14 ± 0.48 | 0.72 ± 0.91 | <.01 |
| Mucous | 4.06 ± 1.97 | 0.29 ± 0.2 | <.01 |
| Ciliatedb | 0 | 0 | |
Source: Ref. 4 Reproduced by permission.
The data for vitamin-A-deprived cells were derived from
epithelia showing minimal changes after five weeks on diet. Foci
of stratification and/or epidermoid metaplasia (about 5% of all
epithelial cells) were excluded from the analysis.
a. Mean percentage of total number of epithelial cells
counted in cross-section of two tracheal rings per hamster (about
1,600 cells). Each hamster received 3H-thymidine and colchicine
six hours before sacrifice.
b. Ciliated cells do not divide.
c. Mean percentage of total number of basal cells or
mucous cells.
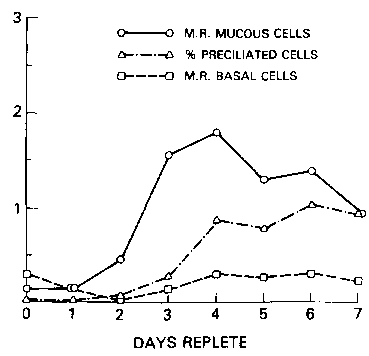
Administration of retinyl acetate to vitamin-A-deficient hamsters, again, primarily affects cell division of the mucous cells (fig. 4). Within three days their cell-division rate returned to normal levels. The number of preciliated cells, which are progeny of replicating mucous cells, was restored to normal levels, but vitamin-A repletion had no effect on the replication of the basal cells.
The first unequivocal conclusion from this in viva work is that vitamin A is necessary to maintain normal rates of mucous cell division in the tracheal epithelium.
In vitro cell culture work
Recent work is] has permitted observation of the effects of vitamin-A depletion in cultured epithelial cells from hamster tracheas. The cells recapitulated the development of normal epithelium during seven days of culture. In the presence of retinoic acid, the mucous cells divided at a high rate and the progeny rapidly matured to fully differentiated mucous and ciliated cells (fig. 5). Smaller cells were also visible in the culture and probably represented basal cells found in viva. The study compared the morphology and cell-division rates of the cells in the presence and absence of retinoic acid. It is clear from figure 6 that deficiency of vitamin A markedly lowered the ability of the larger mucous cells to divide.
When cells grew in the absence of retinoic acid on collagen substrate, they failed to mature into normal columnar mucous cells and, instead, showed the squamous type of differentiation characteristic of vitamin-A deficiency in vivo and of organ-cultured epithelium (fig7).
This in vitro work confirms the notation that retinoic acid in required for cell division and differentation of mucous cells.
The control of mucus production by vitamin A
In conjunction with the lowered rate of mucous cell division, vitamin-A deficiency also causes a decrease in periodic-acid Schiff base (PAS)-positive cells, which indicates less production of mucus. Whether this precedes or follows the decrease in cell division is unclear, but it certainly appears to precede the enhancement of keratin production in the trachea.
Squamous metaplasia is not observed in the small intestine, even in severe vitamin-A deficiency [7]; however, mucin biosynthesis, as measured by the incorporation of 3H-glucosamine, is decreased [8]. We were able to raise an antibody to the purified goblet cell glycoprotein [9]. Indirect immunofluorescence studies clearly indicated the presence of a cross-reactive antigen in a variety of rat epithelial tissues. Vitamin-A deficiency caused a marked drop in the amount of cross-reacting antigen in a variety of epithelial tissues, including the trachea [10]. Therefore, we can conclude that, in addition to cell division of mucous cells, vitamin A also controls mucus production.
Vitamin-A control of keratin gone expression
A later event during the progression of vitamin-A deficiency is keratin gene activation and consequent keratin production. A co-ordinated expression of acidic and basic keratins takes place in vitamin-A deficient hamster tracheas [11]. These keratins are not readily detectable in hamsters fed vitamin A or in tracheas cultured in the presence of retinoic acid. Notwithstanding the morphological similarity between the epidermis and the squamoid epidermoid tracheas, keratin gene products expressed in vitamin-A-deficient tracheas are not the same as in the skin. One outstanding difference is that keratin 1 (67,000 daltons), a prominent epidermal keratin, is not produced in vitamin-A-deficient tracheas [11].
Carcinogenesis
Exposure to chemical carcinogens, such as benzo[a]pyrene and 7,12-dimethylbenz[a]anthracene, causes similar squamous metaplastic lesions in the trachea [12]. When tracheas from four-week-old hamsters fed a normal diet were cultured in the presence of either benzo[a]pyrene or 7,12-dimethylbenz[a]anthracene for two weeks without retinoic acid, they developed squamoid metaplastic lesions. These lesions were not visible when retinoic acid was included in the medium containing the carcinogens (F. L. Huang et al., in preparation). Thus, clearly, retinoic acid is capable of repairing the squamoid metaplasia caused by carcinogen exposure. The squamoid lesions caused by the presence of the carcinogen were also positive by immunofluorescence with keratin antibodies (F. L. Huang et al., in preparation).
Importance of nutritional status in histogenesis: The concept of exotrophism
Organisms capable of synthesizing various essential nutrients are said to be prototrophic for those nutrients. Neurospora is an organism that is prototrophic for pantothenic acid, among other nutrients. When Neurospora is treated with a mutagenic dose of ultraviolet radiation, the result may be the establishment of a requirement for pantothenic acid in the offspring, which is said to have become a "pantothenic-acid auxotroph." Utilizing this approach, Beadle and Tatum [13] have elucidated the steps involved in the biosynthesis of various essential nutrients. Thus, auxotrophism is defined by the need for the exogenous supply of a nutrient in the presence of a mutated phenotype [14].
The mucociliary tracheas, or other epithelia, need vitamin A to maintain their differentiation. The tracheal squamoid cell, which prevails under conditions of vitamin-A deficiency, however, apparently does not need the vitamin. We propose to call this squamous cell "exotrophic" for vitamin A, that is, as having escaped the vitamin-A-requiring status normally characteristic of the columnar epithelium of the trachea. In the presence of vitamin A, this cell is replaced by the normal columnar cells. If, however, the supply of the vitamin is scarce, a conditional "vitamin-A exotroph," that is the squamoid cell, may persist at the site. This exotroph may become permanent by the action of a mutagenic agent, which would fix the exotrophic state, as shown schematically in figure 8. It is then possible to postulate that, when subject to the action of carcinogens and/or tumour-promoting substances, fixed exotrophic cells might divide and contribute to a tumour.
What is the advantage of the exotrophic state to the tumour cell? Quite simply that it has bypassed the requirement for the essential substance and thus the stringency of normal growth control. A cell can become more autonomous as it bypasses the requirements for more essential nutrients. The end result would be a cell that has been liberated from the usual constraints of regulatory substances and that may eventually prevail over other, normally regulated cells.
We and others have examined hepatoma cells and have found that, in general, they contain very little if any retinyl palmitate, whereas the surrounding hepatic host tissue contains normal concentrations of retinyl palmitate [15]. Transplanted hepatomas, whether minimally or maximally deviated from normal, are devoid of retinyl palmitate, whereas the host rat liver tissue and regenerating liver contain normal levels of the vitamin (table 2). Naturally, the lack of detectable retinyl palmitate in hepatoma tissue may be due to a variety of reasons, which are not mutually exclusive, among which are the following:
TABLE 2. Retinyl palmitate of hepatoma, host rat and regenerating liver postnuclear membranes (nanograms per milligram of protein)
| Liver | Tumour | |
| 20-1-1 | 580 ± 30 | <1.6 |
| 16-2-1 | 513 ± 52 | <1.6 |
| 7787-1-1 | 400 ± 21 | <1.6 |
| 9618A-1-1 | 942 ± 2.2 | <1.6 |
| 44-1-2 | 547 ± 21 | <1.6 |
| 5123D-1-1 | 150 ± 3.8 | <1.6 |
| 3924A-1-1 | 177 ± 59 | <1.6 |
| 7800-1-1 | 73 ± 1.8 | <1.6 |
| 5123 tc 1-2 | 363 ± 6.7 | <1.6 |
| 7777-2-1 | 243 ± 38 | <1.6 |
| Primary cystic tumour | 302 ±28 | <1.6 |
| Regenerating liver | ||
| 24 hours | 113 ± 7 | |
| 48 hours | 100 ±10 |
Summary and conclusions
In this paper we have suggested the new concept of exotrophic cells, i.e. cells that have conditionally escaped the need for an essential nutrient, such as vitamin A. These exotrophs might become fixed by a mutation and eventually contribute to the tumorigenic phenotype.
The discovery of the retinoic acid receptor (RAR) has opened up new horizons in the search for the mechanism of action of retinoic acid [17; 18]. It is intriguing that a second retinoic acid receptor, RARE, is abundantly expressed in hepatoma tissue and not in normal liver; Benbrook et al. [191 suggest that the erroneous expression of the RARE might contribute to tumour development in liver. How and whether these findings relate to the vitamin-A-deficient status of hepatoma cells remains to be understood.
References
Michele R. Forman
This paper reviews recent clinical and field research in vitamin A and morbidity and examines, primarily, the epidemiologic evidence for the association between vitamin-A deficiency and morbidity. Since I have been requested to discuss the findings from the most significant work, this review is not exhaustive and is based on an epidemiologist's view of significance. From the available literature, most vitamin-A research focuses on the following morbidities: respiratory disease, diarrhoeal disease, and measles complications, of which respiratory and diarrhoeal infections and corneal ulcers are the most common in less-developed communities. The research was designed as either longitudinal cohort studies of pre-school-aged children or hospital-based trials.
For this review, several criteria were used to evaluate individual studies, including the following:
A summary of study findings and recommendations for future research follow the review of recent research.
Review of recent research
Prospective cohort studies
Prospective cohort studies of vitamin-A deficiency and respiratory and diarrhoeal disease have been con ducted in central Java, Indonesia, and Hyderabad, India. Neither study, however, was designed as a vitamin-A and morbidity study. In an 18-month longitudinal cohort study of pre-school-aged central Javanese children (N = 4,600), ocular and other clinical data were collected at baseline and every three months for evidence of xerophthalmia and respiratory and diarrhoeal disease; anthropometric data were collected to evaluate nutritional status. The child's ocular status was assessed by an ophthalmologist at each three-month examination. The morbidity data were based on clinical signs of current respiratory disease, diagnosed by a paediatrician at the examination, and the diarrhoeal disease data were based on maternal recall of the child's experiencing four or more loose stools per day at any time during the month prior to the examination.
Two separate sets of analyses have been published from this study [1; 2]. In one set [1] the incidence rates of respiratory disease and the prevalence rates of diarrhoeal disease at the end of each three-month interval were determined for children with and without mild xerophthalmia (Bitot's spots and/or night blindness) at both the onset and end of the cycle. Compared with healthy children without xerophthalmia, those children with mild xerophthalmia at the start and end of a three-month cycle had an almost two and threefold risk of respiratory and diarrhoeal disease respectively (table 1). Age-specific relative risks were not appreciably different for diarrhoeal disease and were varied for respiratory disease. These findings were similar within weight-for-height groups (based on the Waterlow classification).
In the second set of analyses [2], all children without xerophthalmia at the beginning of a cycle were stratified by the presence or absence of respiratory or diarrhoeal disease, and the prevalence of mild xerophthalmia was determined at the following examination three months later. Those children with either respiratory or diarrhoeal disease at the onset of a cycle had 2.2 and 2.5 times as great a relative risk respectively of developing mild xerophthalmia by the " end of the cycle as healthy (non-respiratory/non-diarrhoeal disease) controls (table 2). The excess risk of xerophthalmia among respiratory disease cases was restricted to children three years old and over, and the relative risks of xerophthalmia among diarrhoe al cases versus children without diarrhoeal diseases varied by age. These findings were similar within weight-for-height groups.
TABLE 1. Age-specific incidence of respiratory and diarrhoeal disease among children with and without xerophthalmia
| Age (years) | Number of child-intervals | Cases of disease | Rate per 1,000 | Relative risk (N-N: X-X) | p (2-tailed) | |||
| N-N | X-X | N-N | X-X | N-N | X-X | |||
| Respiratory disease | ||||||||
| L 1 | 5,484 | 42 | 470 | 8 | 86 | 191 | 1:2.2 | <.05 |
| 2 | 2 993 | 143 | 257 | 31 | 86 | 217 | 1:2.3 | <.001 |
| 3 | 3 051 | 188 | 176 | 21 | 58 | 112 | 1:1.9 | <.01 |
| 4 | 3,031 | 164 | 100 | 13 | 33 | 79 | 1:2.4 | <.001 |
| 3 5 | 3,644 | 191 | 89 | 5 | 24 | 26 | 1:1.1 | NS |
| Total | 18,203 | 728 | 1,092 | 78 | 60 | 107 | 1:1.8 | <.001 |
| Diarrhoea | ||||||||
| L 1 | 5,425 | 36 | 421 | 9 | 78 | 250 | 1:3.2 | <.001 |
| 2 | 3,014 | 135 | 202 | 31 | 67 | 230 | 1:3.4 | <.001 |
| 3 | 3,018 | 160 | 151 | 27 | 50 | 169 | 1:3.4 | <.001 |
| 4 | 2,958 | 147 | 93 | 14 | 31 | 95 | 1:3.1 | <.001 |
| 3 5 | 3,624 | 183 | 87 | 14 | 24 | 77 | 1:3.2 | <.001 |
| Total | 18,039 | 661 | 954 | 95 | 53 | 144 | 1 :2.7 | <.001 |
Source: Ref. 1.
N-N: Children with normal eyes at both the start and the end
of the three-month observational interval.
X-X: Children with mild xerophthalmia (night blindness and/or
Bitot's spots) at both the start and the end of the interval.
TABLE 2. Age-specific incidence of xerophthalmia for children with and without respiratory disease and for children with and without diarrhoea
| Age (years) | Child-intervals | Developed xerophthalmiaa | Rate per 1,000 | Relative risk
( -: + ) |
|||
| - | + | - | + | - | + | ||
| Respiratory disease | |||||||
| L 1 | 5,533 | 595 | 6 | 0 | 1.1 | 0 | - |
| 2 | 3001 | 417 | 8 | 1 | 2.7 | 2.4 | 1:0.9 |
| 3 | 3061 | 257 | 10 | 3 | 3.3 | 11.7 | 1:3.6 |
| 4 | 3042 | 170 | 11 | 2 | 3.6 | 11.8 | 1:3.3 |
| 3 5 | 3657 | 137 | 13 | 3 | 36 | 219 | 1:6.2 |
| Total | 18,294 | 1,576 | 48 | 9 | 2 6 | 5.7 | 1:2.2b |
| Diarrhoea | |||||||
| L 1 | 4,990 | 465 | 4 | 2 | 0.8 | 4.3 | 1:5.4 |
| 2 | 3038 | 289 | 8 | 1 | 2 6 | 3.5 | 1:1.3 |
| 3 | 3,045 | 172 | 11 | 1 | 3.6 | 5.8 | 1:1.6 |
| 4 | 2979 | 151 | 11 | 2 | 3.7 | 13.2 | 1:3.6 |
| 3 5 | 3644 | 93 | 14 | 2 | 3.8 | 21.5 | 1:5.7 |
| Total | 17,696 | 1,170 | 48 | 8 | 2.7 | 6.8 | 1:2.5b |
Source: Ref. 2.
Minus and plus signs indicate the absence or presence respectively of respiratory disease at the examination initiating a three-month interval, or of a history of four or more loose stools a day within the month preceding the examination.
a. Xerophthalmia present at the examination terminating
the three-month interval.
b. p < .05,2-tailed.
TABLE 3. Incidence of respiratory disease and diarrhoea for children under five years of age with and without mild xerophthalmia-India and Indonesia
Country and age (years) |
Child-intervalsa |
Cases of disease |
Rate per 1,000 |
Relative risk (+ X /-X) |
p (2-tailed) |
|||
-X |
+X |
-X |
+X |
-X |
+X |
|||
Respiratory disease |
||||||||
| India | ||||||||
| L 1 | 1,540 | 3 | 153 | 0 | 99 | 0 | 0.0 | |
| 2 | 724 | 11 | 49 | 3 | 68 | 273 | 4.0 | |
| 3 | 756 | 21 | 36 | 2 | 48 | 95 | 2.0 | |
| 4 | 600 | 27 | 29 | 4 | 48 | 148 | 3.1 | |
| <5 | 3,620 | 62 | 267 | 9 | 74 | 145 | 2.0 | .06 |
| Indonesia | 14,559 | 537 | 1,003 | 73 | 69 | 136 | 2.0 | < 001 |
Diarrhoea |
||||||||
| India | ||||||||
| L 1 | 1.540 | 3 | 961 | 1 | 624 | 333 | 0.5 | |
| 2 | 724 | 11 | 363 | 5 | 501 | 455 | 0.9 | |
| 3 | 756 | 21 | 269 | 11 | 356 | 524 | 1.5 | |
| 4 | 600 | 27 | 175 | 10 | 292 | 370 | 1.3 | |
| <5 | 3,620 | 62 | 1,763 | 27 | 488 | 435 | 0.9 | NS |
| Indonesia | 14,415 | 478 | 867 | 81 | 60 | 169 | 2.8 | <.001 |
Source: Ref. 3.
-X: without xerophthalmia. +X: with xeropthalmia. See text for
definitions.
a. Six-month intervals for India; three-month intervals for
Indonesia.
A prospective community-based study of all preschool-aged children living in the slums of Hyderabad, India (N = 1,544, based on a population registry), was conducted to estimate the incidence of post-measles corneal disease and its relationship to nutritional status [3]. At baseline, all children were examined by medical officers for signs of vitamin-A deficiency and for clinical signs of lower-respiratory infection, diarrhoea, measles, kwashiorkor, and marasmus; weights were also measured. A diarrhoeal-disease episode was defined as a day with three or more loose stools. This baseline examination was repeated at the next two six-month intervals. After the baseline examination, a surveillance system was established whereby field workers determined morbidity status from weekly home visits.
The method of analysis followed that of the central Java data set [1], but classification as vitamin-A deficient for a six-month interval was restricted to the child's xerophthalmia status at the onset of the interval. For children who were mildly xerophthalmic at the onset of a six-month cycle, the risk of lower-respiratory infection, relative to that for non-xerophthalmic children, was 2 (p = .06), and the relative risk of diarrhoeal disease was close to 3 (table 3). These results were not adjusted for potential covariates.
All incidence episodes during a six-month interval were counted in estimating the relative risk. This cumulative episodic approach to the estimation of relative risk and broader definition of a diarrhoeal case than in the central Java study should increase the relative risk for diarrhoeal disease, assuming that mildly vitamin-A-deficient children would have more episodes of diarrhoeal disease during an interval than non-vitamin-A-deficient children. The rates of diarrhoeal disease among the xerophthalmic and non-xerophthalmic were not, however, significantly different, with a relative risk of 0.90.
In sum, the central Java analyses demonstrated a two-way association between vitamin-A deficiency and diarrhoeal or respiratory infection. The Indian analyses supported an association between mild vitamin-A deficiency and an increased risk of respiratory disease, but they did not demonstrate any association between vitamin-A deficiency and diarrhoeal disease. 95% confidence intervals around each relative risk were not provided in the published reports, it is difficult to determine the extent to which the relative risks overlap.
In both studies, children with corneal xerophthalmia were hospitalized, treated with 200,000 IU of vitamin A, and dropped from the study. Children with severe systemic disease were referred to a health facility. These exclusions for ethical reasons may have reduced the relative risks of xerophthalmia and of infection respectively.
Let us examine the ability to detect an association from the data-collection techniques of these studies. In the central Java study, a child's xerophthalmia status was assessed at an examination every three months, and the diarrhoeal data were based on maternal recall for the month preceding the examination. In the data analysis, this cross-sectionally or retrospectively based classification was considered to be stable across the three-month interval. That is, if a child was diagnosed with mild xerophthalmia at the onset and end of a three-month cycle, then he or she was classified as a xerophthalmia if a mother recalled at least one day during the month preceding the exam in which the child had four or more loose stools, then the child was classified as a diarrhoeal-disease case. However, if a mother with a xerophthalmia child selectively recalled more diarrhoeal episodes than mothers of children without mild xerophthalmia, then this systematic bias could alter the estimate of the relative risk. A crucial question for these investigators is indeed whether the mothers of xerophthalmia children recalled more diarrhoeal disease than mothers of children without xerophthalmia. This analysis would include all children in the study (including those with severe systemic disease who were sent to the hospital) rather than only those with or without mild xerophthalmia at both the onset and end of a three-month period.
In the Indian data analysis, a child's xerophthalmia status at the beginning of a six-month period determined her or his classification for that interval. Given the seasonality of vitamin-A-containing foods in many communities and of vitamin-A deficiency 14]. a healthy child could develop xerophthalmia and vice versa over six months. Thus, children could be misclassified among the healthy and among the xerophthalmics. Such bidirectional misclassification of the exposure variable would bias the estimate of the relative risk of morbidity toward the null value and may have indeed done so [5]. With their weekly surveillance data, the investigators might also be able to analyse the potential association between vitamin-A deficiency and severity and duration of respiratory infection, which would be well worth examining.
An additional effort to clarity the association between vitamin-A deficiency and morbidity requires the collection and analysis of data on potential covariates, such as socio-economic status and nutritional status. For example, poverty may be associated with risk of vitamin-A deficiency and of diarrhoeal disease. Adjustment for socio-economic status may appreciably reduce any association between vitamin-A deficiency and respiratory or diarrhoeal disease [6]. Low weight for a given height may be associated with an acute illness [7] and vitamin-A status. In the central Java study, one might want to analyse those ill with diarrhoeal disease at the examination within levels of weight for height to reduce the potential bias from data collected by maternal recall.
In summary, there is a potentially strong crude association between mild xerophthalmia and an increased risk of a new episode of respiratory disease, but few covariates, if any, have been adjusted in the published reports. The association between vitamin-A deficiency and severity of respiratory disease is unknown. The association between vitamin-A deficiency and the risk and severity of diarrhoeal disease is unclear. The directionality of these associations is also unclear. Since the objective of the above-mentioned studies was to examine either blindness or post-measles corneal ulcers, there is a great need for research on vitamin-A deficiency and morbidity.
Clinical trials
Clinical trials conducted in the United States and Australia are noteworthy because of the consistency of their findings regarding the relationship between vitamin-A status and risk of new respiratory infection [8; 9]. In both trials, the investigators presumed that infants and children at high risk of respiratory infection due to lung injury from either neonatal pulmonary insults from hyaline-membrane disease or from lower-respiratory disease early in childhood would benefit from prophylactic doses of vitamin A, as evidence by reduced incidence of subsequent respiratory disease.
The US trial was a randomized, double-blind, hospital-based study of 40 very low birthweight (<1,300 g) neonates to examine the effect of vitamin-A supplementation on the risk of broncho-pulmonary dysplasia (BPD) [hi. At enrolment, the neonates were clinically and anthropometrically comparable, with mean serum retinal levels of 20 fig per decilitre per group. An intramuscular injection of retinyl palmitate (2,000 IU) or of saline solution was administered to each group every other day for 28 days. During the trial, the vitamin-A-supplemented group experienced increased levels of serum retinal and serum-retinol-binding protein (RBP) following the initial dose of vitamin A, but the placebo group did not show a significant change in the serum retinal and RBP levels (fig. 1). Thus, the change in the serum retinal and RBP levels in the vitamin-A group indicated higher rates of mobilization of vitamin A. No evidence of toxicity from hypervitaminosis-A was identified in the vitamin-A group.
The incidence of BPD and clinical signs associated with it were significantly reduced among the vitamin-A-supplemented group as compared with the placebo group at the end of the trial (45% versus 85%, p < .008) (table 4). The number of infants requiring mechanical ventilation by the end of the trial was significantly lower in the vitamin-A group than in the placebo group (21% versus 55%). The vitamin-A-supplemented group required significantly fewer days of intensive care than the placebo group (63 versus 79, p < .02). However, during the first six months of life, four infants in the vitamin-A group and none in the control group died. Probable causes of death were related either to severe hyaline-membrane disease before enrolment in the trial, which led to BPD in an infant who died on day 13 of the trial, or to postnatal infections acquired two or more months after completion of the trial.
The Australian trial was an offshoot of a pneumo caccal vaccine trial (N = 1,273) [8]. Children one to four years old with a history of 15± days of cough or three separate episodes of respiratory illness during the three months preceding the vitamin-A trial were enrolled (N = 47). Children were randomly allocated to a vitamin-A dose of 3,828 IU or a look- and taste-alike placebo without vitamin A. The treatments were administered three times weekly for six months. Mean serum retinal levels at the onset and completion of the trial were 49 and 50 fig per decilitre respectively, in the two groups (table 5). Mothers kept daily records of signs and symptoms of respiratory illness over the trial period.
Children in the vitamin-A supplemented group who received over 50% of the dosage during the trial were included in the analysis. The vitamin-A-supplemented group had approximately 20% fewer respiratory episodes on average than the placebo group (p = .049) (table 6). Among those with a history of acute or chronic lower-respiratory infection (i.e., prior history of bronchitis, pneumonia, croup, whooping cough, or persistent cough) before the trial, the supplemented group had a 25% lower average rate of respiratory disease episodes than the placebo group (p < .05) (table 7).
Thus, in both trials the vitamin-A-supplemented group experienced a significantly reduced risk of a new episode of respiratory infection compared with the placebo group. These trials were conducted among infants and children who were at high risk of severe respiratory infection and who had different vitamin-A status, neonates with marginal serum vitamin-A levels and children with normal serum vitamin-A levels.
A cautionary note about each trial is in order. In the BPD trial, the increased mortality among the vitamin-A-supplemented group is problematic. An four deaths were attributed to BPD. One possible explanation for this problem focuses on uncontrollable post-trial factors, such as distance from the subject's home to a health-care facility. Although this explanation is conjectural, future research should, as suggested by the authors, maintain surveillance of the diet, morbidity, and vitamin-A status of the study subjects beyond the end of the trial.
The findings of the Australian trial are quite exciting, but because of methodological shortcomings they must be viewed with caution. Notably, the analysis compared the rates of respiratory episodes among the placebo group and those who had a 50% or more compliance with the treatment schedule among the vitamin-A group. Such an analysis destroys the randomization component of the study design. The investigators need to reanalyse their data using all subjects in the vitamin-A group or adjust the current respiratory infection rates of the vitamin-A group to those factors distinguishing the non-compliants from the compliants. Moreover, it is not clear why there was a 20% difference in the rates of respiratory episodes between the vitamin-A and placebo groups but a much smaller, insignificant difference in the mean number of days with specific respiratory symptoms. Perhaps the investigators can describe the missing data about the respiratory-disease episodes that determine the significant difference between the supplemented group and the placebo group. Finally, given the small cell sizes, it may always be uncertain whether these findings are real or due to chance, and, therefore, the trial requires replication.
TABLE 4. Clinical outcome of infants during
| Vitamin-A supplemented | Control | p | |
| Incidence of broncho pulmonary dysplasia | |||
| N | 9/20 | 17/20 | |
| % | 45 | 85 | <.008 |
| Need for mechanical ventilation on study day 28a | |||
| N | 4/19 | 11/20 | |
| % | 21 | 55 | <.029 |
| Ventilatory requirements on study day 28a | |||
| FiO2 | 783 ± 381 | 895 ± 241 | <.040 |
| Ventilator rate | 223 ± 530 | 313 ± 338 | NS |
| peak inspiratory pressure | 112 ± 224 | 231 ± 215 | NS |
| positive end-expiratory pressure | 29 ± 43 | 66 ± 49 | <.020 |
| mean airway pressure | 59 ± 107 | 101 ± 76 | <.040 |
| oxygenation index | 0.63 ± 0.36 | 0.69 ± 0.15 | < 030 |
| Sepsis | |||
| episodes per infantb | 2.5 ± 1.0 | 3.1 ± 1.7 | NS |
| airway infectionc | |||
| N | 4/19 | 11/20 | |
| % | 21 | 55 | <.029 |
| Retinopathy of prematurity | |||
| N | 5/19 | 12/20 | <.034 |
| % | 26 | 60 |
Source: Ref. 9.
Plus/minus values are mean ±SD. NS: not significant.
a. Area-under-curve values obtained by plotting multiple
readings of variable in 24-hour period against time. Patient 1,
who died on postnatal day 13 is excluded from this analysis.
b. Clinically suspected sepsis with or without
confirmation by microbiologic culture resulting in initiation of
antimicrobial therapy.
c. Confirmed by positive microbiologic cultures of airway
secretions.
Table 5. Mean plasma vitamin-A concentrations in supplemented and unsupplemented groups
| Retinol (1g 100ml) | |||||
| Initial | Final | ||||
| Mean | SE M | Mean | SE M | N | |
| Placebo | 48.9 | 2.0 | 49.9 | 1.6 | 43 |
| Supplement | 50.5 | 1.8 | 50.2 | 1.7 | 47 |
Source: Ref. 8.
In sum, compared with the placebo group, the vitamin-A-supplemented group had a reduced incidence of new episodes of (BPD) respiratory disease. The Australian trial data were stratified by history of allergy and of lower-respiratory infection, but no other adjustments for potential covariates were made. These trials were conducted among infants and children at high risk of respiratory disease, but this effect has not been demonstrated among pre-school-aged children who are not at risk of severe respiratory infections.
Vitamin-A deficiency and measles complications
Now let us turn to the potential relationship between vitamin-A deficiency and measles complications, including respiratory and diarhoeal disease, corneal ulcers, and mortality. Measles is a highly contagious disease that infects all susceptibles when exposed. Infection with this virus is characterized by impaired host immunity, which may increase the risk of complications among the already malnourished [10; 11]. Measles reduces energy intake, utilization, and absorption and damages the epithelial cells of the respiratory and gastrointestinal tracts [12; 13]. All these factors may act individually or in combination to increase the risk of complications.
TABLE 6. Mean respiratory symptoms in vitamin-A-supplemented and placebo groups
Placebo (N=54) |
Supplement (N= 53) |
Difference (%) |
Significance (p)a |
|
Mean SE M |
Mean SE M |
|||
| Episodes of all symptoms | 8.0 0.57 |
6.5 0.45 |
-19 | .049 |
| Days of all symptoms | 72.7 5.7 |
72.7 8.7 |
0 | NS |
| Nose (days) | 62.5 5.2 |
54.7 7.4 |
-12 | NS |
| Cough (days) | 28.3 3.6 |
32.2 5.3 |
-12 | NS |
| Chest (days) | 15.1 3.3 |
13.7 5.0 |
-9 | NS |
Source: Ref. 8.
a. Student's t-test. NS: not significant.
TABLE 7. Percentage differences between supplemented- (S) and placebo-group (P) means (P - S) stratified by allergy and lowerrespiratory-illness (LRI) history
| History of allergy | History of LRI | Total population | |||
| No | Yes | No | Yes | ||
| N | 63 | 44 | 39 | 68 | 107 |
| Episodes of all symptoms | -11 (NS) | -25 (NS) | 0 (NS) | -25(P < .05) | - 19 (P < .05) |
| Days of all symptoms | 17 (NS) | -16 (NS) | 4 (NS) | -1 (NS) | 0 |
Source: Ref. 8.
Vitamin A has an important role in cell reproduction and differentiation of the respiratory and gastrointestinal tracts, mucosa of other organs, immune system, and conjunctiva/cornea of the eye [14-21]. Given the physiological functions of vitamin A, its deficiency has the potential to act synergistically with measles infection in the incidence of measles complications. Indeed, measles and vitamin-A deficiency have similar complications, such as diarrhoea, acute respiratory disease, and xerophthalmia [20; 22-27].
There are two recent publications on this subject. In the 15-month community-based study of post-measles complications and nutritional status in a slum area of Hyderabad, India, that was discussed earlier [3], the measles attack rate among pre-school-aged children was 23% (N = 318/1,544) [28]. Among the measles cases, the incidence rate for bronchopneumonia was 34%, for diarrhoea 37%, and for corneal lesions 3% (table 8). Children between one and two years of age suffered from the highest rates of measles and measles complications.
TABLE 8. Incidence of complications during measles
| Age (years) | No. of measles cases | With broncho- pneumonia | With diarrhoea | With corneal lesions |
| <1 | 69 | 24 | 29 | 3 |
| 1+ | 105 | 40 | 45 | 4 |
| 2+ | 62 | 21 | 22 | 0 |
| 3+ | 47 | 16 | 11 | 0 |
| 4+ | 35 | 8 | 11 | 3 |
| 0-4+ | 318 | 109 | 118 | 10 |
| (100%) | (34 3%) | (37.1%) | (3 1%) |
Source: Ref. 28.
The mean serum retinal levels of the Indian children were always below 20 fig per decilitre regardless of measles status (table 9). During measles, the mean serum retinal levels of children with and without secondary infections or corneal lesions were not significantly different (table 10). During fever, serum retinal levels are spuriously lowered, and in a small subsample of 32 measles cases the serum retinal levels were lower during the acute phase than during the pre- and post-measles period (table 11). Malnourished measles cases (based on Gomez classification) had significantly lower mean serum retinol and albumin levels than their better nourished counter-parts (table 12).
In a Tanzanian randomized trial of hospitalized measles cases who received either a high dose of vitamin A (200,000 IU) on two consecutive days (N=88) or standard therapy without vitamin a (N=92), the case fatality rate during the month following measles was 7% (N=6/88) in the vitamin-A-supplemented group (p=.13) (table 13) [29]. A significant difference was observed among children under two years old, with a 2 % versus 17% case fatality rate (p<.05). Additionally, marasmic children had a fourfold higher case fatality rate than better nourished children regardless of treatment allocation. These findings should be considered suggestive because the total number of deaths (N=18) was small and trial was not double-blind inasmuch as the paedriatricians knew each subject’s treatment group.
TABLE 9. Biochemical parameters (cross-sectional data)
| Serum levels | ||||
| N | Albumin (g/dL) |
Retinol (µg/dL) |
RBp (mg/dL) |
|
| Control (C) | 117 | 3.6 ± 0.03 | 17.9 ± 0.81 | 2.5 ± 0.06 |
| Measles during infection | ||||
| (Ma) | 153 | 3.4 ± 0.03 | 11.5 ± 0.44 | 2.1 ± 0.07 |
| after recovery (M2) | 108 | 3.5 ± 0.05 | 19.8 ± 0.69 | 2.5 ± 0.06 |
Source: Ref. 28. Values are mean ± SE
Statistical significance -
C vs. M1 p < .001 for albumin for retinol and for
RBP.
M1 vs. M2: NS for albumin; p< .001 for
RBP.
TABLE 10. Biochemical parameters (longitudinal data on TABLE 13. Mortality of children admitted with measles 32 children)
| Serum levels | |||
| Albumin (g/dL) | Retinol (µg/dL) | RBP (mg/dL) | |
| Premeasles (PM) | 3.6 ± 0.07 | 16.5 ± 1.75 | 2.4 ± 0.14 |
| Measles ( M, ) | 3.4 ± 0.08 | 11.1 ± 1.1 | 2.2 ± 0.17 |
| Postmeasles (M2) | 3.7 ± 0.07 | 19.2 ± 1.68 | 2.3 ± 0.15 |
Source: Ref. 28.
Statistical significance -
PM vs. M1: p<.05 for albumin; p< .02 for
retinal; NS for RBP.
M1 vs. M2: p < .01 for albumin, p <
.001 for retinol: NS for RBP
TABLE 11. Serum vitamin-A levels in relation to measles and corneal lesions
| Corneal changes in measles | N | Albumin (g/dL) |
Retinol (µg/dL) |
| No change | 47 | 3.3 ± 0.09 | 13.7 ± 1.37 |
| Coarse keratitis | 19 | 3.2 ± 0.09 | 10.8 ± 1.08 |
| Fine keratitis | 6 | 3.2 ± 0.14 | 11.6 ± 1.39 |
| Corneal xerosis | 20 | 3.3 ± 0.07 | 11.5 ± 1.63 |
Source: Ref. 28.
TABLE 12. Biochemical parameters in measles according to nutritional status
| Nutrition Grade | No. of children | Albumin (g/dL) | Retinol (µg/dL) | RBP (mg/dL) |
| Normal+ | ||||
| Grade I | 58 | 3.4 ± 0.05 | 12.5 ± 0.80 | 2.0 ± 0.80 |
| Grade II | 74 | 3.4 ± 0.05 | 11.3 ± 0.60 | 2.2 ± 0.09 |
| Grade III | 21 | 3.2 ± 0.07a | 8.3 ±0.75b | 2.2 ± 0.15 |
Source: Ref. 28.
a. p < .05 compared with Normal + grade I.
b. p < .02 compared with Normal + grade I.
TABLE 13. Mortality of children admitted with measles
Age |
Children admitted | Children who died | ||
| Given vitamin A |
Controls | Given vitamin A |
Controls | |
| <9 | 14 | 9 | 0 | 2(22) |
| 9 - 11 | 12 | 10 | 0 | 2(20) |
| 12-23 | 20 | 23 | 1(5) | 3(13) |
| 24-35 | 11 | 16 | 3(27) | 2(13) |
| 36-47 | 11 | 13 | 1(9) | 1(8) |
| 48- 59 | 8 | 6 | 1(13) | 0 |
| ³ | 12 | 15 | 0 | 2(13) |
| Total | 88 | 92 | 6(7) | 12(13) |
Source: Ref. 29.
Principle data are numbers of children. Figures in parentheses
are percentages.
TABLE 14. Measles complications and associated mortality
Complication |
Children with complications |
Children who died |
||
Given vitamin A |
Controls |
Given vitamin A |
Controls |
|
| Pneumonia | 38 (43) |
47 (51) |
3 (8) |
7 (15) |
| Otitis media | 19 (22) |
20 (22) |
1 (5) |
3 (15) |
| Group or laryngo tracheobronchitis | 8 (9) |
13 (14) |
1 (50) |
4 (31) |
| Dysentery | 2 (2) |
6 (7) |
1 (4) |
3 (50) |
| Haemorrhagic rash | 28 (32) |
34 (37) |
1 (11) |
4 (12) |
| Oral candidiasis | 9 (10) |
5 (5) |
1 (20) |
|
Source: Ref. 29.
Principle data are numbers of children. Figures in parentheses
are percentages.
Based on the two measles studies presented above, vitamin-A deficiency and measles complications do co-occur but are not necessarily associated. Nevertheless, the pathophysiology is sufficiently similar in measles and vitamin-A deficiency to suggest the need for future research in this area.
The evidence from the hospital-based trial leads us back to the potential association between vitamin-A deficiency and respiratory disease inasmuch as respiratory-specific mortalities were reduced among the vitamin-A-supplemented versus the control group (table 14). Given the findings of respiratory-disease-specific mortality differences in the Tanzanian trial, a follow-up study might be designed as a randomized double-blind trial of vitamin-A supplementation (versus a placebo) administered to measles and non-measles respiratory-disease cases to compare their risk of mortality.
A curious feature of the Tanzanian trial is the absence of any diarrhoeal-disease deaths. In the Indian study [28|, 34% of the measles cases suffered from diarrhoeal-disease complications during the acute stage of measles, and 76% suffered from diarrhoeal disease during the six months following measles. Thus, the clear absence of any diarrhoeal deaths in the Tanzanian trial is noteworthy.
Before leaving the subject of vitamin-A deficiency and measles complications, I would like to discuss current World Health Organization recommendations 130] briefly. The WHO and the United Nations Children's Fund recommend that a high-potency preparation of 100,000 and 200,000 IU of vitamin A be administered to measles cases under and above one year old respectively. They state that this therapeutic regimen should be followed in communities with measles fatality rates of 1% or higher or in communities with recognized vitamin-A deficiency. This recommendation is based on the above-mentioned hospital-based trial in Tanzania, but the dosages differ. Moreover, this recommendation is presented for a broader range of communities than can be extrapolated from the trial: vitamin-A distribution to measles cases is recommended for any community with a measles-case fatality rate of 1% or more regardless of its vitamin-A status.
Before such a recommendation is implemented, several studies might be conducted, including a trial of vitamin-A supplementation (versus a placebo) before the measles season to examine the risk of the incidence and duration of measles complications. Another trial might examine the impact of a high-potency preparation of vitamin A (versus a placebo) administered to non-hospitalized measles cases during the acute stage on the incidence and duration of complications. These trials would address the preventive effect of the administration of vitamin A compared with that of a placebo prior to and during the measles season.
Summary and research recommendations
Let me briefly summarize this review and discuss several issues regarding future research. First, there is a potentially strong association between vitamin-A status and the risk of respiratory infection. Second, it is more difficult to state the potential for the association between vitamin-A deficiency and diarrhoeal disease based on the techniques of morbidity assessment or the analytic approach to data taken in published articles. It is noteworthy that the studies presented were not designed to examine the association between vitamin-A deficiency and morbidity. Third, vitamin-A deficiency and measles complications co-occur, and indeed, vitamin-A supplementation during measles may reduce respiratory-specific causes of mortality. These statements refer primarily to crude associations that require adjustment for potential covariates in future research.
One cannot address future research in vitamin-A deficiency without examining methods of vitamin-A assessment and detection of morbidity. Essentially, researchers must be able to determine subclinical and clinical vitamin-A deficiency in order (a) to compute an unbiased estimate of the risk of morbidity among these two groups in comparison with those who have adequate vitamin-A status and (b) to examine the potential threshold effect of vitamin-A status on morbidity. Various methods of vitamin-A assessment exist, but ideally they should be evaluated for their field applicability [31; 321, their sensitivity and specificity across the spectrum of vitamin-A deficiency, and their reproducibility before research on vitamin-A deficiency and morbidity is conducted.
Another complicating issue in vitamin-A assessment is determining the vitamin-A status of febrile children, notably children with acute infections, because fever spuriously lowers serum retinal levels. This issue needs to be addressed in virtually all vitamin-A and morbidity studies that assess the vitamin-A status at the time of diagnosis of an infection. One plausible method of correcting for the effect of fever on serum retinal levels is to measure the levels of serum acute-phase reactant protein, which is inversely associated with serum retinal levels under stressful conditions (such as after surgery). Thus, the changes in C-reactive protein levels [33] can be used to adjust the serum retinal levels to pre-infection status.
The ability to detect an association between vitamin-A deficiency and diarrhoeal and respiratory infections, as well as the direction of that association, is contingent on the quality of morbidity data collection. The information required for diagnosis and determination of the severity of infection includes standardized clinical validation and, preferably, microbiologic analysis. In a proposed University of Gadjah Mada and Johns Hopkins University vitamin-A and morbidity field trial [34], my colleagues and I are developing a morbidity surveillance system of twice-weekly home contact with the child's caretaker in order to identify symptoms and signs of infection. Within 24 hours of the identification of a potentially ill child, a physician will *sit the home to validate the diagnosis clinically and take a specimen for microbiologic analysis. With this information, we will be able to distinguish single from multiple episodes, the duration of each episode, and acute from chronic episodes. With such data one can examine whether vitamin-A deficiency increases the risk of a new episode and/or the severity of an infection and, indeed, the association between vitamin-A deficiency and etiologic-specific agents of diarrhoeal and/or respiratory disease.
If future studies of vitamin A and morbidity are designed as clinical of field trials, then some attempt should be made to validate the proper dosage of vitamin A to be administered in the trials. Two hundred thousand IU of vitamin A is the high-potency preparation of vitamin A that will be administered in all the ongoing mortality trials and the proposed morbidity trials. This dosage is based on one study of Brazilian children under 10 years old who had a relative-dose-response test at baseline, followed by the administration of a dose of 200,000 IU of vitamin A. At 30, 120, and 180 days after the dosing, the relative-dose-response test was repeated to determine the time interval until return to the baseline serum retinal levels 1351. The impact of dosages lower than 200,000 IU on the risk of morbidity or mortality has not been examined, but the ethical objectives of current field trials should be the administration of the lowest dosage of vitamin A that potentially reduces the risk of morbidity.
All research on vitamin A and morbidity should be conducted in communities with evidence of clinical and/or subclinical vitamin-A deficiency to ensure that there will be beneficial effects of a trial by treating children with xerophthalmia. Thus, recent data on vitamin-A deficiency rates within the past five years are required before the onset of the trial.
Finally, covariates that may potentially influence the association between vitamin A and morbidity include season; dietary intake of vitamin A, protein, and fats; anthropometric status, including weight and height; intestinal parasites that may reduce the ability to absorb vitamin A; socio-economic status; access to and utilization of health care; and child's age and sex. Information about these variables should be collected during a study, and the variables should be examined as potential interaction terms, effect modifiers, and/or confounding variables with respect to the association between vitamin-A deficiency and morbidity.
References
{kind=link}
{kind=link}
{kind=link}
{kind=link}